Introduction
FIRST CHAPTER - EXPERIMENTAL ESTABLISHMENT OF THE STATE OF HABITUATION
First chapter
PART ONE - CRITICAL REVIEW OF PREVIOUS STUDIES
I. Animals used
PART TWO - SELECTION OF AN ANIMAL CAPABLE OF MORPHINE HABITUATION
II. Habituation of Rats
Concentrations of the solutions of morphine hydrochloride in relation to doses injected
Weights of three rats before the beginning of the experiment and after seven weeks of regular injections
Average duration* of sleep induced by intravenous injection of 25 δ per g. of sodium evipan, in rats which have been given one or two successive subcutaneous injections of morphine hydrochloride
Average duration* of sleep (minutes) induced by the intravenous injection of 25 δ per g. of sodium evipan in habituated rats
Determination of the toxicity of heroin administered by intravenous injection
Concentration of solutions and mortality in relation to doses of heroin injected
Determination of the toxicity of heroin administered by subcutaneous injection
Determination of the toxicity of heroin administered by intravenous injection
Average duration* of sleep (min.) caused by intravenous injection of 25 δ per g. of sodium evipan in rats having received two successive subcutaneous injections of heroin
Author: Denise G. Fichtenberg
Pages: 19 to 42
Creation Date: 1951/01/01
The Bulletin hereafter presents the first part of an original Work, fruit of many years' research and experiments on the very controversial question of habituation to morphine. The author, a Doctor of Natural Sciences, is chief of biological work at the Laboratory of Physiological Control of Anti-venereal Medicaments (French Ministry of Public Health), and has achieved experimentally the state of habituation in the rat, has worked out techniques of biological assay for morphine, studied the fixation of morphine in the organism and its elimination, and discovered that the organism of habituated animals transforms or destroys more morphine than that of normal animals.
It has long been known that the human organism can tolerate usually toxic doses of certain poisons without grave consequences, provided those doses are administered regularly. This acquired tolerance has been given the name of mithridatism or habituation.
In a man Who takes morphine daily, the symptoms are characteristic and complex.
As a result of repeated administration of the drug, the usual doses cease to have their elective effect upon many individuals To; cause analgesia, increasingly larger doses, even in excess of normally toxic quantities, have to be used. This principal phenomenon is in most cases accompanied by an overwhelming craving for the drug, and the withholding of the latter causes phenomena known as disorders of abstinence or deprivation.
Since the phenomena of deprivation or abstinence characteristic in the morphine addict are not normally found in morphinized animals or with the majority of poisons giving rise to habituation, a number of authors (Jeanne Levy, 1934; R. Cahen, 1935) distinguish, at least for the purpose of experimental study of the mechanisms of habituation, between habituation proper and abstinence phenomena.
The Bibliography of the article will be found at the end of the second part in the next issue of the Bulletin.
Nevertheless, in the case of morphine, a theory of the mechanism of habituation to this substance has to explain both the weakening of analgesic action and the tolerance of toxic doses which are characteristic of habituation proper, as well as the craving for the drug.
The state of habituation has aroused the curiosity of many experimenters who have tried in vain to discover more about its mechanism. They have worked along different lines and followed different conceptions; their results are often contradictory. No valid explanation of habituation has been found so far.
Much work has been done to determine by comparative methods what becomes of morphine in the organisms of normal and habituated animals and how it is eliminated. So far, however no proof has been adduced of more rapid or more complete disappearance of morphine in the case of habituated animals. It should be noted that the divergencies observed by various authors can be explained: (1) by the lack of precision with which the degree of habituation in tested animals has been determined in most cases; (2) by the difficulty of extracting; describing and assaying morphine in tissues, biological liquids and excretions.
In this study I have tried to verify two theories advanced in the past, namely that: (1) morphine becomes transformed into its oxydation product, oxydimorphine; (2) morphine circulates in the blood in a less concentrated form in the organisms of habituated animals than in those of normal animals.
Before undertaking a study of what happens to morphine in the organism of animals, I had to resolve two fundamental problems: (1) that of finding an animal showing habituation phenomena, upon which a series of experiments could be carried out; (2) that of establishing a specific and sufficiently precise method of assaying morphine, oxydimorphine and morphine accompanied by oxydimorphine.
Having at my disposal an animal capable of habituation-the rat-and a pharmacological test permitting the detection of 0.030 mg. of morphine accompanied by oxydimorphine, I worked out a simple method of extracting morphine from tissues and biological liquids, and studied the fixation of morphine and its conjugated form in various organs and its elimination. The purpose of this work was to measure the quantity of morphine administered and determine what percentage of morphine is converted within the organism into a substance devoid of analgesic effect.
The following plan was used in preparing the study:
Part one: Critical review of previous studies
I Animals used.
II Tests employed to control the state of habituation.
III Summary and criticism.
Part two: Selection of an animal capable of morphine habituation
I Effect of repeated morphine injections upon mice.
II Habituation of rats:
(A) Tolerance of toxic doses of morphine hydrochloride;
(B) Physical condition of rats habituated to morphine;
(C) Diminution of analgesic action during habituation;
(D) De-habituation;
(E) Reaction to hypnotics of rats habituated to morphine;
(F) Habituation to heroin;
(G) Summary.
SECOND CHAPTER - BIOLOGICAL ASSAYS OF MORPHINE?
I Catatony of the mouse's tail.
II Action of cholinic esters On the denervated muscle of the leech under the influence of morphine:
(A) Study of the sensitizing action of morphine on the acetylcholinic contraction of the denervated muscle of the leech; Establishment of a biological assay of morphine;
(B) Study of the antagonist effect of morphine on the contraction caused by various cholinic esters in the denervated muscle of the leech; Establishment of a biological assay of morphine and oxydimorphine.
THIRD CHAPTER. WHAT HAPPENS TO MORPHINE IN THE ORGANISM
Part one: Detection of oxydimorphine
I Action of blood on morphine in vitro.
II Detection of oxydimorphine in vivo.
Part two: Disappearance, conjugation and accumulation of morphine in vitro by tissues and in vivo in normal and habituated rats
I Action of certain tissues upon morphine.
II Content of free and conjugated morphine in the blood and tissues of normal and habituated rats.
Part three: Elimination of morphine
I Elimination of free and conjugated morphine through the urine and faeces of normal rats.
II Elimination of free and conjugated morphine through the urine and faeces of various groups of habituated rats.
Part four: Comparison between quantities of morphine accumulated in the organism or eliminated by normal and habituated rats
CONCLUSIONS
Experimental Establishment of the State of Habituation
In the first part of this chapter I shall relate briefly and critically the work of various experimenters who have tried to build up tolerance experimentally,* then in the second part I shall describe my personal research.
After reviewing the various animals which were experimented upon, I shall examine the various tests used by those conducting the experiments to establish the degree of habituation of animals subjected to morphine.
The animals most frequently used are the dog,1 the rabbit2 and the rat.3
* In a very complete survey Eddy has studied tolerance and habituation in all animal species.
1Faust (1905); Van Egmond (1911); Myers (1916-1939); Takayanaki (1934); Tatum, Seevers and Collins (1927, 1929); Pierce and Plant (1927, 1930, 1932); Barbour, Hunter and Richey (1929); Hotta (1932); Birk (1933); Giordano (1933); Koh (1933); Schmidt and Livingston (1933); Wolff, Riegel and Fry (1933); Cahen (1934-1935); Hayashi (1934); Ikeshima (1934); Anton and Birk (1935); Plant and Slaughter (1936-1938); Donnini (1937); Gross, Plant and Thompson (1938); Wada, Tanaka, Hirano and Taneiti (1936); Barbour, Porter and Seelye (1939); Gross and Thompson (1940); Thompson and Gross (1941); Gross (1942); Slaughter, Treadwell and Gales (1943); Wright and Sabine (1943); de Bodo (1944); Goetz, Burril and Ivry (1944); Shideman (1946); Templeton, Calapeaux and Adler (1940); etc.
2Cloetta (1903); Marie (1907); Van Egmond (1911); Takayanaki (1924); Gottlieb (1926); Tatum, Seevers and Collins (1929); Sue (1932); Koh (1933); Cahen (1934, 1935, 1936) Hayashi (1934); Ro (1934); Ko (1935); Shou (1935); Keil and Pohls (1936); Donnini (1937); Puru (1937); Hinohara (1937); Kuwahara (1937-1938); Delaville (1938); Emerson and Phatak (1938); Endo (1938); Ingersoll (1940); Okawa (1940); Sato (1940); Tin (1940); Yosikawa (1940); Ra (1940); Wright and Sabine (1943); etc.
3Rubsamen (1908); Hildebrandt (1922); Takayanaki (1924); Joel and Ettinger (1926); Ma (1932, 1933, 1936, 1937, 1939); Cheng and Ma (1934); Hayashi (1934); Cross and Pierce (1935) Himmelsbach, Gerlach and Stanton (1935); Weger and Amsler (1936); Amsler (1937); Donnini (1937); Fitshugh (1939); Seevers and Shideman (1941); Dietrich and Thienes (1941); Shideman and Seevers (1941, 1942); Lewis (1949); etc.
The literature on the subject also contains accounts of a considerable number of experiments per formed on the guinea-pig,4 the mouse,5 and in more exceptional cases the cat6 and the monkey.7
In these animals habituation was realized through the injection of morphine over varying periods (several days, several months, several years) by various methods (intramuscular, subcutaneous, intraperitoneal, buccal); the morphine is administered either in small constant doses (Gross and Thompson, 1940; Goetz, Burril and Ivry, 1944, with dogs), or in progressively increasing doses (Ma, 1932, with rats) or in rather large doses from the outset (Schmidt and Livingston, 1933, with dogs; Rubsamen, 1908, with rats).
II.Tests employed to control the state of habituation
To control the degree of habituation of animals experimented upon, the authors resorted to one or more of the four following tests:
Diminution of the analgesic effect produced by the active initial dose.
Tolerance to toxic doses for normal animals.
Disappearance of secondary actions brought on by morphine.
Particular reaction of animals habituated to morphine to the active initial dose of various substances (cross habituation).
On the basis of the definition of hahituation given in the introduction, the first two tests used simultaneously may be considered as characteristic of that state provided they are applied to a sufficient number of animals. On the other hand the application of the other two tests to animals habituated to morphine has not been studied sufficiently systematically to make it possible to determine the degree of habituation of animals on the basis of these two tests alone.
Many authors have not experimented systematically and simultaneously with the first two tests which are characteristic of habituation and have merely verified the weakening of the analgesic action or the various secondary effects of morphine.
4Amsler (1931); Fleischman (1931); Keeser, Oelkers and Raetz (1933); Hayashi (1934); Bertschik (1934); Weger and Amsler (1936); Donnini (1937); Matschulan (1937); Delaville and Russel (1938); Rentz and Kesarbani (1941); Frommel, Piquet, Cuehod, Loufti and Aron (1945).
5Kreitmair (1928); Keeser, Oelkers and Raetz (1933); Koh (1933); Hayashi (1934); Cahen (1935); Favarget and Rosselet (1944).
6Cold (1929); Tatum; Seevers and Collins (1929); Hotta (1931); Hayashi (1934).
7Tatum, Seevers and Collins (1929); Abe (1936); Hotta (1931); Seevers (1934); Yokota (1935).
To give only a few examples, we may mention Rubsamen (1908) who notes the tolerance of the rat to morphine as soon as he can absorb food, half an hour after the injection; Himmelsbach, Gerlach and Stanton (1935), who evaluate the degree of habituation of the rat by his greater or lesser irritability, when he is placed in an abnormal position after the injection; Kobayashi (1930), who observed a diminution in the narcotic effect and in myosis in the dog and rabbit while the rhythm of the heart, respiratory frequency and vagal action are not modified; Tatum, Seevers and Collins (1929), who observed in dogs habituated to morphine a weakening in the analgesic and vomitive action, a greater tolerance of heroin, codeine, chloralhydrate and barbiturates without any simultaneous material variation in the minimum lethal dose.
The divergent and sometimes contradictory observations hitherto noted are no doubt due to this lack of homogeneity and precision.
We shall examine below the results obtained by using these various tests.
Reduction of the analgesic effect produced by the active initial dose.
Few authors have systematically studied the wearing off of the analgesic effect brought about by the regular administration of morphine. Nevertheless it has been shown by a variety of tests that in dogs, tolerance to analgesia is not established, while that tolerance is manifest in guinea-pigs and rats.
Goetz, Burril and Ivry (1944), who gave daily subcutaneous injections of 3 mg. of sulphate of morphine per kg. to dogs for fifty days, measured the threshold of painful excitement produced on the dental pulp by electric stimulation. The thresholds of painful sensations were measured twice a week, firstly before the injection of morphine and secondly thirty, sixty and ninety minutes after the injection. The threshold of pain after rising, decreases following the daily injections, only to increase again with the effect tending to vary periodically. The authors conclude that during the period of daily administration of a uniform dose of morphine, the analgesic action decreases but the tolerance is not lasting and disappears and re-appears periodically.
Weger and Amsler (1936) determined in the normal guinea-pig, and every, two days during the daily administration of morphine, the number of stimuli required to produce a squeal after subcutaneous injection of 50 mg. of morphine hydrochloride. The analgesic action decreased in the course of the experiments.
Rentz and Kesarbani (1941) made subcutaneous injections of increasing doses of morphine hydrochloride, varying from 25 to 100 mg. per kg. from sixty to eighty days on guinea-pigs. As from the twenty-first day the animal was no longer sensitive to the painful sensation observed with the active initial dose in the normal guinea-pig.
Lewis (1949), using the Ercoli and Lewis test (1945) (reaction to thermic stimulus) in rats, measured the degree of analgesia caused by morphine, heroin, codeine, dilaudide and demerol (dolosal). In particular in one group of twenty rats which received subcutaneous daily injections of 10 mg. of sulphate of morphine per kg. for six weeks, the analgesic effect decreased regularly until it reached 40 per cent of its original force.
Tolerance to toxic doses for normal animals
To establish this tolerance, the minimum lethal dose must be determined accurately. The majority of the research workers who have studied the mechanics of habituation have failed to determine it with sufficient precision. Nevertheless, experiments made on various animal species show on the one hand that in the dog, the guinea-pig, the rabbit and the mouse, there is no tolerance to doses which would be toxic for normal animals, whereas a certain tolerance is built up in the rat. Thus Rubsamen (1908) and Hildebrandt (1922) were able to make subcutaneous injections in the last-mentioned animal of doses between 710 mg. per kg. and 1,000 mg. per kg., doses which are known to be much higher than the toxic dose for the normal animal.
Dog:
Tatum, Seevers and Collins (1929), experimenting on twenty dogs, indicated that. the toxic dose varies from 200 to 416 mg. per kg. for normal animals and from 250 to 416 mg. per kg. for animals having received subcutaneously 3 mg. per kg. of sulphate of morphine daily for six months. Schmidt and Livingston (1933) observed the development of tolerance to a dose of 100 mg. per kg. in one month in dogs which received 30 to 60 mg. per kg. daily, three-and-a-half to four months in dogs receiving 2 to 10 mg. per kg. daily. The mortality rate was much higher in the first case than in the second.
Cahen (1935) did not work on a sufficiently large number of animals to enable him to determine the habituation of the dog to morphine. He injected dogs subcutaneously every two days for two-and-a-half to three months with progressively increasing doses from 1 to 35 mg. per kg. and obtained the results indicated in the table given below:
Determination of the minimum lethal dose for normal and habituated dogs (according to Cahen, 1935)
|
Normal dog |
Habituated dog | |||
|---|---|---|---|---|
|
Dose of (HCl) in mg/kg |
No. of animals experimented |
No. of death |
No. of animals experimented |
No. of death |
| 100 | 2 | 0 | 2 | 0 |
| 135 | 5 | 5 | 4 | 1 |
| 150 | 2 | 1 | 5 | 2 |
| 200 |
- |
- |
2 | 1 |
Templeton, Calapeaux and Adler (1940) merely observed that a dose of 162 mg. of sulphate of morphine injected intravenously produces the same rate of mortality in normal dogs and those habituated to morphine.
Rabbit:
Experiments made on rabbits are not any more precise. It should be noted indeed that toxicity in rabbits varies considerably with age, according to Eddy (1939) who experimented on 3,000 young rabbits. The difference of toxicity according to age may explain the contradictory results obtained by the different authors referred to.
Tatum, Seevers and Collins (1929) found that after prolonged administration of morphine, this animal cannot bear a minimum lethal dose greater than the dose tolerated by the normal adult animal, 100 to 200 mg. per kg.
Cahen (1935), who administered morphine hydrochloride to rabbits for three months by subcutaneous injection, in increasing doses varying from 1 to 50 mg. per kg. notes that a dose of 200 mg. per kg. injected intravenously causes the death of five out of six rabbits while the ratio is four out of six in normal animals.
On the other hand, Furu (1937) pointed out that the rabbit tolerates large doses of morphine after repeated injections of heroin and codeine. It is probably a matter of cross-habituation.
Guinea-pig:
Cahen (1935) experimented with guinea-pigs after regular administration of morphine. Every two days, the animals received subcutaneous injections of morphine hydrochloride in doses varying from 0.4 to 200 mg. per kg. The toxicity created subcutaneously had the following results:
Determination of the minimum lethal dose for normal and habituated guinea-pigs (according to Cahen, 1935)
|
Normal animal |
Habituated animal | |||
|---|---|---|---|---|
|
Dose in mg/kg |
No. of animals |
No. of death |
No. of animals |
No. of death |
| 500 | 7 | 3 |
- |
- |
| 550 | 6 | 6 | 7 | 5 |
Rat:
The experiments made with rats were not conducted with sufficient attention to detail especially as Cheu and Robbins (1944), studying the toxicity of sulphate of morphine injected intravenously, found that young rats were more sensitive than older animals, and the age of the animals is not mentioned in the experiments made by Rubsamen (1908). and Hildebrandt (1922). Moreover, Rubsamen did not systematically determine the minimum lethal dose for normal animals and animals habituated to morphine. He merely noted that 510 to 600 mg. per kg. of morphine hydrochloride injected subcutaneously was toxic for the normal animal. By administering 300 mg. per kg. to a group of rats, he obtained a mortality of about 50 per cent. He injected doses increasing gradually up to 710 mg. per kg. in the survivors and observed a reduction in mortality. Hildebrandt (1922) habituated young rats during their period of growth to subcutaneous injections of morphine. The doses are increased slowly from 100 to 600 and 1,000 mg. per kg.
Mouse:
Cahen (1935) injected increasing doses of morphine hydrochloride from 10 to 60 mg. per kg. into mice sub- cutaneously daily for two-and-a-half months and obtained the following results:
Determination of the minimum lethal dose for normal and habituated mice (according to Cahen, 1935)
|
Normal animal |
Habituated animal | |||
|---|---|---|---|---|
|
Dose in mg/kg |
No. of animals injected |
No. of death |
No. of animals injected |
No. of death |
| 620 | 6 | 3 |
- |
- |
| 650 | 6 | 6 | 6 | 2 |
The mouse habituated to morphine thus tolerates a dose only slightly larger than that tolerated by the normal animal.
Disappearance of the secondary effects of morphine
Many research workers have assumed that an animal becomes accustomed, to morphine when there is a change in the Various secondary effects of the drug: sedative action; action on thermogenesis, action on respiration, on the Cardiac vagus, on the pupil, on vomiting, on the gastro-intestinal tract and on the genital organs.
The observations of the various authors do not always concord. The over-all results obtained show that the variations in the secondary effects of morphine in the habituated animal are not simultaneous: the sedative action is the first to disappear, the cardiac vagus becoming habituated later. The divergences noted in connexion with the secondary effects, may arise from the method of habituation and from the species of animal subjected to experiment, as will be seen from the description which follows:
Narcotic and sedative action. A normal animal injected with morphine falls in to a drowsy state for a varying length of time. Repeated doses of the drug reduce the length of the period or eliminate it. The authors observed that the state of drowsiness is characterized particularly by a change in posture reflexes or reflexes of the lower limbs, and the animal's irritability after the injection. They noted that the monkey, dog, rat and mouse withstand the sedative effect of morphine.
In the monkey: Hotta (1931) and Seevers (1934) observed a weakening of the narcotic effect of morphine beginning on the third day of regular injection of the drug.
In the dog: Plant and Pierce (1927) note only a reduction in the duration of the state of drowsiness as the animal becomes accustomed to the drug.
Tatum, Seevers and Collins (1929), by regularly injecting 3 mg. per kg. in the animal obtain no more than a slow weakening of the state of drowsiness. On the other hand, Kobayashi (1930) reports that a dose of 60 mg. puts a normal dog to sleep while 2 g. in the same animal, once he is habituated leaves him awake after the injection.
Wilker and Frank (1948) show that in the spinal dog the reflexes causing the flexing and extending of the hind leg become weaker after a single dose of morphine and return to normal with regular injections of the drug.
According to Kobayashi (1930) the guinea-pig does not become accustomed to the sedative effects of morphine.
Himmelsbach, Gerlach and Stanton (1935) noted irritability in the rat when tied to a board and flagellated. In its normal state, this animal becomes insensible after an injection of morphine. After eight days of daily injections of morphine, the animal struggles under the influence of the initial dose.
Cahen (1935) notes slight drowsiness in the normal mouse which precedes catatony of the tail; in the mouse habituated to morphine, he observes only the catatonic state without drowsiness.
Action on thermogenesis. Morphine lowers the temperature of the normal animal. This action has been studied during habituation in the rabbit; the results obtained are contradictory.
While Girndt and Lipchitz (1931) find no habituation to thermal morphine reactions by chronic intoxication of the animal, Cahen (1935) notes a rise to more than one degree above normal in the temperature of the rabbit habituated to morphine for three months, and Ko (1937) points out that the hypothermy induced by morphine in the normal animal disappears with repeated injections of the drug.
Action on respiration. In the normal animal, morphine has a depressive effect on respiration which takes the form of a great diminution in the frequency and an increasing in the depth of respiratory movements.
The disappearance of the respiratory action due to morphine, after repeated and prolonged administration of the drug, has not been noted in all cases in different animals, nor by all authors in the same animal.
In monkeys Hotta (1931) states that a few daily injections of morphine eliminate the cyanosis engendered by a single injection of morphine in a normal animal.
In dogs habituated to morphine, Van Dongen (1915), Schmidt and Livingston (1929), Hotta (1931) and Cahen (1935) report the disappearance of the inhibitory effect of morphine on the respiration; on the other hand, Tatum, Seevers and Collins (1929) noted no distinct changes in the respiratory response after prolonged administration, for four months, of 3 mg. per kg.; these results are definitely confirmed by Kobayashi (1930), who reports the same slowing down of respiratory frequency after injections of morphine in normal dogs and dogs habituated to the drug.
Hotta (1931) reports that in cats no habituation of the respiratory centre was observed after forty-three daily injections of morphine into those animals.
Cahen (1935) and Okawa (1940) observed that the depressive action of morphine on the respiratory system of the rabbit did not change throughout the prolonged administration of the drug.
Action on the cardiac vagus. Morphine gives rise to bradycardia in normal animals by stimulating the vagal centre. This stimulation, principally observed in dogs, does not change in animals habituated to morphine, according to Tatum, Seevers and Collins (1929) and Kobayashi (1930), or is diminished slightly, according to Hotta (1931), by the regular administration of morphine for twenty to twenty-two months.
Pupillary reaction. An injection of morphine gives rise to myotic action in monkeys, dogs and rabbits. Hotta (1931) noted that myosis in these animals disappeared on the repeated administration of morphine. The same phenomenon had already been reported by Van Dongen (1915) and Kobayashi (1930) in dogs.
In normal cats, morphine brings about mydriasis, which persists after prolonged administration of the drug (Hotta, 1931).
Vomitory action. The generally acknowledged emetic effect of morphine on normal dogs is questioned by Tatum, Seevers and Collins (1929) and Cahen (1935), who state that a large dose of morphine administered immediately has no emetic effect on a normal animal.
The vomitory action of the drug is said to be less in dogs habituated to morphine (Van Dongen, 1915; Downs and Eddy, 1928; Tatum, Seevers and Collins. 1929; Cahen, 1935), and in cats (Hotta, 1931). Nevertheless, according to Hotta (1931), this action persists in dogs habituated to morphine.
Castro-intestinal phenomena. (α) In situ. In normal animals, morphine gives rise to a decrease in the tonicity, frequency and extent of stomachic movements, to an increase in the tonicity and peristaltic movements of the small intestine and of the ileo-caecal sphincter and to an increase in the tonicity of the rectum.
The effect of morphine on the gastro-intestinal tract is not greatly changed by the repeated administration of the drug. Van Egmond (1911) showed that the depressive action of morphine on the stomach persists in dogs habituated to morphine. Miller and Plant (1926) and Templeton, Calapeaux and Adler (1940) found that daily injections of morphine into dogs continued to produce a marked increase in the tonicity, extent and frequency of intestinal peristaltic movements. On the other hand, Cahen (1935) observed a decrease in the intestinal effects of morphine on dogs habituated to morphine.
A morphine injection usually produces a diarrhoeic effect on a dog. Kobayashi (1930) and Hotta (1931) found that this effect continued in an animal chronically under the influence of morphine; Du Mez (1919), Plant and Pierce (1928), Schmidt and Livingston (1929), and Tatum, Seevers and Collins (1929), observed a decrease in defaecation and a tendency to constipation in animals habituated to morphine.
( β) Isolated organ. The tonicity of the isolated intestine of a normal animal is reduced when it is placed in contact with a solution of morphine. The phenomenon is reversed in the case of the isolated intestinal loop of an animal which has received morphine regularly. This effect was observed by Kobayashi (1930) in the isolated intestines of the dog and rabbit and by Jeanne Levy and Cahen (1933) in the isolated intestines of the rabbit and guinea-pig.
Action on the genital organs (α) Development of the genital organs. The regular administration of morphine prevents the normal development of the male or female genital organs, as was observed by San Fillipo (1939) in young dogs which had received the drug regularly for twelve months. The genital organs suffered fatty degeneration and the ovaries contained no mature follicles. Morphinism in mice disturbs the genital cycle (Ko, 1939).
( β) Isolated organ. Morphine causes increased tonicity if it is placed in contact with the isolated uterus of a normal rabbit. Kobayashi (1930) observed the contrary reaction of the uterus to morphine in a rabbit which received daily injections of the drug; Ko (1939) observed only a decrease in tonicity.
Special behaviour of animals habituated to morphine on receiving the active initial dose of various substances (cross-habituation )
Research workers have thought it interesting to study the action of depressive or stimulating poisons on animals habituated to morphine and to base the proof of habituation to morphine on these findings.
Jeanne Levy (1934) has made a very complete summary of these studies and I shall only quote the most recent ones, which are confined, to opium derivatives, phenanthrenic alkaloids, cocaine, apomorphine and hyperthermics. There has been no recent research on the effect of barbiturates on animals habituated to morphine.
Opium derivatives and phenanthrenic alkaloids. Rabbits habituated to morphine can tolerate doses of opium alkaloids, phenanthrenic alkaloids or their mixtures: pantopon, laudanum, narcophine, dilaudide, dicodide, and heroin, which are usually fatal to normal animals (Keil and Poels, 1936). The hyperglycemic effect on these animals of heroin, codeine, dionine and pantopon is also less (Ko, 1935).
Cocaine. The local analgesic effect of cocaine on the cornea of a rabbit is increased, both in intensity and duration, after a preliminary injection of morphine. Bertschik (1934) and Amsler (1935) observed a reversal of this phenomenon in rabbits habituated to morphine; cocaine no longer acted as a local anesthetic.
Apomorphine. The vomitory action of apomorphine is reduced in dogs habituated to morphine. Issekutz and Varady (1934) used this action as a test of the existence of habituation in dogs.
Hyperthermics. Plant and Slaughter (1936) found that the buccal administration of 5 mg. per kg. of dinitrophenol (a hyperthermic with peripheral action) in a bicarbonate solution, which gives rise to heightened metabolism in normal dogs, had no effect on animals habituated to morphine. This result was confirmed in rabbits by Cahen (1935), who also found that hyperthermics with central and peripheral action (brewers' yeast, tetrahydronaphthylamine) give rise to greater hyperthermy in animals chronically intoxicated with morphine.
III. Summary and criticisms
Dogs, rabbits, guinea-pigs, rats and mice are the animals most generally used for the study of experimental habituation to morphine. Their behaviour during the prolonged administration of the drug has given rise to varying or contradictory comments.
The authors have not ascertained with sufficient care the existence of habituation in any given animal. They observed differences in the behaviour of the functions and organs of animals which received the drug regularly.
The most typical phenomena of habituation to morphine, tolerance of doses which would be toxic to a normal animal and the lessening or disappearance of the analgesic effects of morphine, have not been experimented on systematically and the results obtained have not been confirmed by all research workers. A toxic dose may be determined by different methods. At present, there is a tendency to determine the dose which is lethal in 50 per cent of cases; this can have no meaning, however, unless the experiment is conducted on a sufficient number of animals of the same species, sex and weight, uniformly fed throughout the experiment and kept at a uniform temperature
The analgesic effects of morphine can be determined by means of different tests on the same animal; but in all cases the experiment should be carried out under absolutely identical conditions on normal animals and on animals receiving morphine regularly.
The results obtained by various research workers, who have not always operated under the above conditions, lead to the following conclusions:
In dogs, habituation to analgesic effects is not manifest; although the toxicity has not been determined accurately, this animal does not tolerate doses greater than those which are toxic to normal animals. The habituation of dogs to morphine was assumed when the secondary effects of the drug were found to have disappeared, each experimenter studying a few specific symptoms. These secondary effects did not all disappear at the same time: the sedative effects diminished first, whereas the vagal action continued for a long time and the effect of morphine on intestinal movements was unchanged. It therefore seems difficult to define the degree of habituation of a dog, which is very small and differs greatly from that observed in human beings.
Rabbits. After regular administration of morphine, they cannot tolerate doses which are toxic for normal animals. Most of the secondary effects of morphine remain in animals which are chronically intoxicated. Rabbits cannot therefore be used for the experimental study of habituation.
Guinea-pigs. The doses tolerated by them are only slightly greater than those which are generally toxic for a normal animal; here again, experiments in toxicity are extremely incomplete; the analgesic effects of morphine diminish during the period of daily administration, but the secondary effects of morphine remain. The guinea-pig is not, therefore, the most suitable animal.
Rats. According to Rubsamen (1908) and Hildebrandt (1922), they tolerate approximately three times the 50 per cent lethal dose for normal animals and, unlike other research workers, the above authors administered these large doses to animals for considerable periods; nevertheless, neither the disappearance of analgesic effects,8 nor the changes in the secondary effects of morphine have been studied systematically and concurrently.
Mice. In spite of certain somewhat unconvincing experiments, these animals were used by Cahen (1935) in experiments in connexion with the study of habituation.
Having at my disposal a selected stock of mice and rats, I carried out experiments on these two species of animals.
I found that the two main tests, the simultaneous application of which makes it possible to demonstrate with accuracy an animal's habituation to morphine, were the following:
(1) Tolerance to doses which would be lethal for a normal animal;
8Lewis (1949) made a comparative study of the analgesic action of morphine and of certain synthetic analgesic substances.
(2) Diminution or disappearance of the analgesic effects caused by the administration of the active initial dose of morphine; reappearance of those effects with the administration of larger doses.
Tolerance to lethal doses can easily be demonstrated by the determination of doses 50 per cent lethal before and after repeated administration of the drug. It is, however, very important to take into account the method by which the poison is introduced in to the organism. When administered by intravenous injection, morphine reaches the organs much more quickly than when administered by subcutaneous or intramuscular injection, and may on that account undergo less complete transformation in the blood and muscles. If that were so, a habituated animal would only be able to tolerate by intravenous injection a dose slightly larger than that tolerated by normal animal, whereas it would tolerate a much larger dose administered by subcutaneous injection.
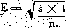
Moreover, Jeanne Levy (1945) has stressed the significance of the ratio toxicity by subcutaneous injection/toxicity by intravenous injection. This is generally about 3(2.5 to 3.5) for poisons which are eliminated unchanged or which only undergo slow transformation in the organism. It may reach higher figures for poisons which are rapidly transformed in the organism into substances less toxic than themselves, while it is less than 2 for poisons which are rapidly transformed into substances more toxic than themselves. The value of this ratio might have led to the discovery of data on the changes undergone by a substance in the organism, and I made a systematic study of it.
Furthermore, since according to some authors morphine is liable to be transformed in vivo into oxydimorphine, I also determined the toxicties of that substance in the normal animal when administered by subcutaneous intravenous injections.
My first research was carried out on the mouse. Since that animal showed no tolerance to 50 per cent lethal doses of morphine administered by subcutaneous or intravenous injection after daily treatment for two months, I gave up the mouse and turned to the rat which, when habituated to morphine for a certain length of time, has a definite tolerance to doses which would be lethal for the normal animal. I carried out a number of experiments on the rat which made it possible to establish the degree of habituation to morphine. I studied in succession the attenuation of the analgesic action of morphine in the course of habituation, and the behavior of a habituated rat to hypnotics (evipan, soneryl). Finally, I studied the rat's habituation to another analgesic, heroin.
I. Effect of Repeated Morphine Injections upon Mice
The first experiments were carried out on the mouse (D. Fichtenberg, 1946) in order to verify whether that animal showed a certain degree of tolerance to morphine during the regular administration of the drug. The animals used, which were about five weeks old and whose growth had been regular, weighed 16 to 19 g. at the beginning of the experiment.
Determination of the 50 per cent lethal dose of morphine hydrochloride when administered by subcutaneous and intravenous injection
Subcutaneous injection. Group of ten or twenty mice were given an injection of a 2 per cent solution of morphine hydrochloride in the flank. The animals, kept at a constant temperature (21°C.), were examined after twenty-four hours and the mortality percentage was noted.
|
Dose (δ per g.)* |
Number of animals |
Mortality percentage |
| 350 | 10 | 10 |
| 450 | 10 | 10 |
| 500 | 10 | 30 |
| 550 | 20 | 40 |
| 580 | 20 | 50 |
| 600 | 10 | 60 |
| 700 | 20 | 70 |
To calculate the average individual variation for a 50 per cent lethal dose, the toxicity curve is plotted on Bonet-Maury logarithm probability paper. The ordinates are the mortality percentages (projection of the S curve along a straight line), and the abscissae are the logarithms of the doses administered.
In the above example (Graph I on page 26) the different points obtained make it possible to draw a straight line which passes approximately through all the points.
The average variation for the 50 per cent lethal dose is calculated by applying the formula where t is the mortality percentage s the survival percentage n the number of animals utilised
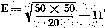
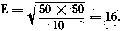
When marked on the toxicity graph ± 11 corresponds to-doses of 540 and 610 respectively. The 50 per cent lethal dose being 580, the difference is: -40 and + 30, with a maximum of +40.
The percentage error is 40 X 100 = 6.9.
The 50 per cent lethal dose of morphine hydrochloride when administered to the mouse by subcutaneous injection is 580 ± 40 per g.
(I) If ten animals are used instead of twenty:
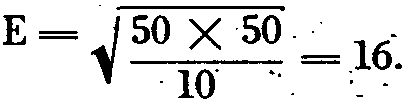
In this article, ? = microgram.
Intravenous injection. Groups of ten or twenty mice were injected in a vein of the tail with a 2 per cent solution of morphine hydrochloride in physiological serum. They were kept at a temperature of 21°C. and examined after twenty-four hours and the percentage established.
|
Dose (δ per g.) |
Number of animals |
Mortality percentage |
| 200 | 10 | 10 |
| 225 | 10 | 20 |
| 250 | 20 | 40 |
| 260 | 20 | 50 |
| 300 | 20 | 90 |
Average variation for the 50 per cent lethal dose: II (Graph II, page 26).
50 per cent lethal dose: 260 ± 20 δ g.
Percentage of error: 7.6 per cent.
Toxicity by subcutaneous injection/toxicity by intravenous injection. For morphine hydrochloride this ratio is 2.2, which, according to Jeanne Levy (1945), would appear to indicate that morphine is not transformed or undergoes very slow transformation in the mouse's organism.
Determination of the 50 per cent lethal dose of oxydimorphine hydrochloride administered to the mouse by subcutaneous and intravenous injection
I used oxydimorphine hydrochloride prepared in the following way:
Preparation of oxydimorphine. I used the Fulton (1933) method with some changes. 2.5 g. of morphine hydrochloride are dissolved in 50 c.c. of water. The base is precipitated with 5 c.c. of standard soda, and the alkalinity of the solution is checked after vigorous shaking. A 1 per cent solution of potassium ferricyanide is added gradually and the liquid is shaken up by electricity for one hour. The presence of an excess of ferricyanide is checked by persistence of the yellow colouring of the solution. The liquid is centrifugalized and decanted and the precipitate is washed twice with distilled water. The precipitate is dissolved over heat in N/5 hydrochloric acid. To obtain complete transformation of the morphine into oxydimorphine, it is preferable to filter and to repeat the same operations again on the filtrate. The final precipitate, dissolved in hydrochloric acid, is brought to a 6.5 pH by means of concentrated ammonia. It is centrifugalized and the decanted liquid is checked to see that it has a pH of 6.5, and-that a few drops of dilute ammonia do not make the top liquid turn milky. The latter operation is repeated a second time to purify the oxydimorphine. The final precipitate is washed twice with warm distilled water, which dissolves the mineral salts, and then once with pure alcohol and once with ether. It is then dried in the oven at 37o C.
To check that the product obtained is free from any trace of morphine, Marquis's reagent is used. A small quantity of alkaloid is powdered on a watch glass with a small drop of 12 volume hydrogen peroxide. A drop of sulphuric acid with formalin added to the watch glass produces an emerald-green colouring characteristic of oxydimorphine, whereas traces of morphine give a violet tint spoiling the green coloration.
The base is transformed into hydrochloride by dissolving it in 0.2 N hydrochloric acid and it is evaporated over a water bath to obtain the hydrochloride in the dry state.
Toxicity by subcutaneous injection. Groups of ten mice were injected in the flank with a 2 per cent solution of oxydimorphine hydrochloride. After twenty-four hours the following mortality percentages were obtained:
|
Dose (δ per g.) |
Number of animals |
Mortality percentage |
| 200 | 10 | 0 |
| 500 | 10 | 0 |
| 700 | 10 | 0 |
| 1,000 | 10 | 0 |
| 1,500 | 10 | 35 |
| 2,000 | 10 | 65 |
Average variation for the 50 per cent dose: 16.
50 per cent lethal dose: 1,750 ± 10 δ per g.
Percentage of error: 0.57 per cent.
Toxicity by intravenous injection. The mice, having been injected in a vein of the tail with a 2 per cent solution of oxydimorphine hydrochloride in physiological serum, were examined after twenty-four hours.
|
Dose (δ per. g.) |
Number of animals |
Mortality percentage |
| 50 | 10 | 0 |
| 60 | 10 | 0 |
| 70 | 10 | 0 |
| 80 | 10 | 100 |
| 90 | 10 | 100 |
| 100 | 10 | 100 |
Average variation for the 50 per cent lethal dose: 16.
50 per cent lethal dose when administered by intravenous injection: 75 ± 2.5 δ per g.
Percentage of error: 3.3 per cent.
Toxicity by subcutaneous injection/toxicity by intravenous injection. For oxydimorphine hydrochloride this ratio is 23.3. This substance is sharply differentiated from morphine by the 50 per cent lethal dose being very much greater when administered by subcutaneous injection and very much smaller when administered by intravenous injection. The high value of the ratio R indicates, according to Jeanne Levy (1945), that it must be rapidly transformed in the mouse's organism into a substance less toxic than itself.
Behaviour of mice subjected to daily injections of morphine hydrochloride
The difference in the reaction of the mouse to morphine and to oxydimorphine is sufficient to determine whether the morphine is transformed more completely into oxydimorphine in habituated animals than in the normal animal.
I sought to habituate mice to morphine and determined in these animals the toxicity of this substance after subcutaneous and intravenous injections, as well as the ratio: toxicity by subcutaneous injection/toxicity by intravenous injection.
The mice were treated every second day for two months by means of subcutaneous injections of increasing doses of morphine hydrochloride. The injections are made into each flank alternately; the flanks are cleansed with a 60 per cent alcohol solution both before and after injection. The animals are kept at an even temperature (21°C.).
The morphine hydrochloride is dissolved in distilled water; the concentrations are determined on the basis of an injection of 0.50 c.c. of liquid for a mouse weighing 20 g.
The quantities injected are in proportion to the weight of the animals, which are weighed before injection. The initial dose is equal to 0.050 mg. per g. of body weight. During subsequent weeks the doses are increased from 0.025 mg. per g. twice a week to. 0.275 mg. per g. at the beginning of the eighth week.
A total of 124 male mice were used in the experiment, the mortality rate being about 10 to 12 per cent per week. The catatony of the tail caused by morphine is the same at the beginning, and at the end of the experiment, during which the physical aspect of the mice remains normal.
Determination of 50 per cent lethal doses of morphine hydrochloride
Fifty-one mice survived after eight weeks. Toxicity in these animals is determined by the subcutaneous and intravenous injection.
Subcutaneous injection. The mice are injected in the flank with 1 per cent solutions of morphine hydrochloride, and are examined twenty-four hours after the injection.
|
Dose (δ per g.) |
Number of animals |
Mortality percentage |
| 550 | 10 | 0 |
| 600 | 10 | 60 |
The 50 per cent lethal dose of morphine hydrochloride administered by subcutaneous injection is about 0.590 mg. per g.
Intravenous injection. A solution of 1 per cent morphine hydrochloride in physiological serum was in jected into a vein in the tail of the mice. They were examined twenty-four hours after the injection.
|
Dose (δ per g.) |
Number of animals |
Mortality percentage |
| 200 | 5 | 0 |
| 250 |
l0 |
100 |
| 300 | 8 | 100 |
The 50 per cent lethal dose of morphine hydrochloride administered by intravenous injection amounts to approximately 0.225 mg. per g.
Toxicity by subcutaneous injection/toxicity by intravenous injection. The 50 per cent lethal doses determined after regular injection with morphine hydrochloride are absolutely comparable with those determined in the normal animal.
The ratio of toxicities by these two methods is equal to 2.6, which is not far from that obtained in normal mice (2.2).
Morphine thus does not appear to undergo any speedy change in the organisms of mice subjected to repeated injections of the drug. The mice, after a period of two months, are unable to tolerate doses, whether subcutaneous or intravenous, in excess of the toxic dose for normal mice. Cahen (1935) noted, from experiments on a Small number of mice, a very low rate of tolerance in habituated mice, the mortality rate of which was 34 per cent as against 100 per cent.
Summary
As mice habituated to morphine show no tolerance of doses which are toxic for normal animals, they cannot be used in a study of the mechanism of habituation. I accordingly made no further experiments with this animal.
As stated earlier, the authors who experimented with rats did not determine with sufficient precision the degree of habituation which may be attained by this animal.9 Thanks to homogeneity, selective breeding and the large number of animals placed at my disposal, I was able to undertake a series of experiments to determine the degree of habituation of animals to which morphine had been administered daily by various methods.
A. Tolerance of toxic doses of morphine hydrochloride
This study, parallel to that undertaken with mice, consists in determining the toxicities of morphine and of oxydimorphine in the normal rat, according to the method of injection; and in the daily injection of in creasing doses of morphine and the determination of the toxicities of morphine in rats habituated to the drug. (Fichtenberg, D. G., 1948). A study is also made of the speed of habituation of the rat subjected for a period of three days to twelve weeks to constant doses or to doses increased at more or less rapid rates.
9As this chapter was in the course of preparation we received Lewis' article (1949) which determines with precision the development of tolerance in the rat to the analgesic effect of morphine.
The rats utilized, whether males or non-pregnant females, three months old, have a regular rate of growth and weigh at least 20 g. at one month, 40 g. at two months and 75 g. to 80 g. at three months.
Determination of the 50 per cent lethal dose of morphine hydrochloride administered to normal rats by subcutaneous or intravenous injection
Subcutaneous injection. The animals are injected in the flank with a 4 per cent solution of morphine hydrochloride. They are kept at an even temperature (21 0C.) and examined after twenty-four hours. Preliminary toxicity tests are made on groups of four rats, and the 50 per cent lethal dose is determined on twenty rats (ten males and ten females). The results are as follows:
|
Dose (δ per g.) |
Number of animals |
Mortality percentage |
| 200 | 4 | 0 |
| 225 | 4 | 25 |
| 250 | 10 males | 50 |
| 250 | 10 females | 50 |
| 275 | 4 | 75 |
| 300 | 4 | 100 |
There is no difference in toxicity between males and females. Average Variation for the 50 per cent lethal dose: 11 (Graph IV, page 26). 50 per cent lethal dose when administered by subcutaneous injection: 250 ± 10 per g. Percentage of error: 4 per cent .
Intravenous injection. Preliminary experiments were also undertaken on groups of four rats. A 2 per cent solution of morphine hydrochloride in physiological serum is injected into the internal saphena of a hind foot. The rats are examined after twenty-four hours.10
The following mortality rates were observed:
|
Dose (δ per g.). |
Number of animals |
Mortality percentage |
| 40 | 4 | 0 |
| 50 | 4 | 0 |
| 70 | 4 | 0 |
| 80 | 4 | 0 |
| 90 | 4 | 0 |
| 95 | 5 | 20 |
| 100 | 10 males | 50 |
| 100 | 10 females | 50 |
| 110 | 4 | 100 |
Average percentage for the 50 per cent lethal dose:
11. 50 per cent lethal dose of morphine hydrochloride administered by intravenous injection, identical in males and females: 100 ± 3 δ per g.
10Animals injected by this method die Within two to six hours. None were found to die beyond that period.
Percentage of error: 3 per cent (Graph III page 26).
Toxicity by subcutaneous injection/toxicity by intravenous injection. The ratio for morphine hydrochloride is 2.5. It seems to show that morphine is converted very slowly in the organism of rats as in that of mice.
Determination of the 50 per cent lethal dose of oxydimorphine hydrochloride administered to rats by subcutaneous and intravenous injection
Since hydrochloride is only slightly soluble in water, the quantities of oxydimorphine, to be injected obliged me to prepare benzoic sulphonate (Leuller and Pomme, 1930). This salt is obtained by the action of two molecules of sulphonic acid of benzene on a basic molecule in a solution of water with a slight acid excess; it is concentrated and allowed to crystallize. The product is washed with alcohol which eliminates the excess of sulphobenzoic acid. For the sake of consistency I have expressed the results in oxydimorphine hydrochloride.
Subcutaneous injection. The strength of the solution of oxydimorphine benzoic sulphonate, expressed in oxydimorphine hydrochloride, is 3.62 per cent. The injections are made in the flanks, and the percentages of mortality, after twenty-four hours, are as follows:
|
Dose (δ per g.) |
Number of animals |
Mortality percentage |
| 362 | 4 | 20 |
| 579 | 4 | 30 |
| 640 | 10 | 50 |
| 724 | 10 | 100 |
Average variation for the 50 per cent lethal dose: 16.
50 per cent lethal dose of oxydimorphine hydrochloride administered by subcutaneous injection: 640±90 δ per g.
Percentage of error: 14 per cent.
Intravenous injection. The strength of the solution of oxydimorphine benzoic sulphonate expressed in hydrochloride is 1.8 per cent in physiological serum. The injection is made in the internal saphena of a hind foot, and the rats are examined after twenty-four hours. The percentages of mortality are as follows:
|
Dose (δ per g.) |
Number of animals |
Mortality percentage |
| 24 | 3 | 0 |
| 30 | 4 | 0 |
| 45 | 4 | 0 |
| 91 | 10 | 30 |
| 108 | 10 | 60 |
Average variation for the 50 per cent lethal dose: 16. 50 per cent lethal dose administered by intravenous injection: 102 ± 14 per g.
Percentage of error: 14 per cent.
Toxicity by subcutaneous injection/toxicity by intravenous injection. This ratio for oxydimorphine hydrochloride is 6.5. Although weaker than it is with mice, it is greater than that found for morphine. According to Jeanne Levy (1945), oxydimorphine is converted more rapidly than morphine in the rat's organism into a substance less toxic than itself. The 50 per cent lethal doses of morphine and oxydimorphine are identical When injected intravenously, but different when injected subcutaneously, oxydimorphine being less toxic than morphine.
Daily injection of morphine in rats
The animals are placed four or five in a group in glass crystallizers 250 mm. in diameter covered by a lid with a grill. The glass bowls are cleaned twice a week and strewn with wood shavings. The food, changed every day, consists of Cereal grains (wheat, barley, oats) moistened bread and, twice a week, boiled potatoes. A 150 c.c. flask filled with water is fixed to the lid, the narrow neck enabling the rats to take in the water directly drop by drop. The bowls are placed in a ventilated room kept at a constant temperature of 21oC. The rats used, males and non-pregnant females, are three to three-and-a-half months old and weigh from 75 to 110 g. The flanks of the animals are shaved before the beginning of the experiment, the subcutaneous injections are made in each flank alternately, and they are washed in 60o alcohol before and after the injection.
First experiment: Twenty-one rats received increasing doses of morphine hydrochloride every day (except Sunday) for seven weeks.11
Doses. I chose as an initial dose a quantity equivalent to 40 per cent of the 50 per cent lethal dose for subcutaneous injection: 0.100 mg. per g. That quantity of morphine does not cause death in normal rats. I increased that dose by 0.100 mg. per g., or 4 per cent of the 50 per Cent lethal dose, daily for seven weeks. At the end of the seventh week, the quantity injected amounted to 0.500 mg. per g.: or 200 per cent of the 50 per cent lethal dose.
The concentrations of the solutions varied in terms of the dose, in accordance with the proportions set forth in the table below:
|
Number of weeks |
Doses (in δ per g.) at the beginning of each week |
Concentrations of the solutions (Percentages) |
| 1 | 100 | 1 |
| 2 | 160 | 2 |
| 3 | 220 | 2 |
| 4 | 280 | 3 |
| 5 | 340 | 4 |
| 6 | 400 | 4 |
| 7 | 460 | 4 |
In the remainder of the experiment these animals will be designated as group III.
Behaviour of the animals. The rats become drowsy half an hour after the injection of morphine. This state, which lasts two or three hours, is observed during the first three days of the experiment. On the fourth and fifth days, the animals, although not drowsy, nevertheless find difficulty in standing. This state appears twenty to thirty minutes after the injection and lasts for an hour. From the second week onwards, the injection no longer affects their behaviour, although the quantity of morphine injected is 60 per cent larger than the initial quantity. Thus, there is habituation to the depressive action of morphine.
Growth of the animals. The daily administration of morphine prevents the regular growth of the rats. The average weight of a three-month-old rat is 85 g.; at five months the adult animal weighs an average of 120 g. The weight of the rats subjected to the experiment varies little during the seven weeks of daily injections with morphine. A rat weighing 85 g. at the beginning of the experiment weighs 91 g. two months afterwards. These results are clearly shown in the graph below:
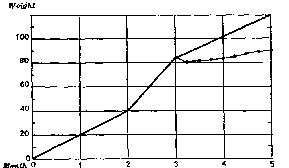
Graph of the average growth of normal rats and of those receiving daily injections of morphine hydrochloride from the third month onwards. From the third to the fifth month, the points show the average weight of the animals subject to the experiment each week.
Mortality during the experiment. The total mortality was eleven rats out of twenty-one, or about 50 per cent. The animals died regularly from the second week of the experiment onwards, as the following table shows:
|
Number of weeks |
Doses in δ per g. injected at the end of each week |
No. of deaths each week |
| 1 | 150 | 0 |
| 2 | 210 | 1 |
| 3 | 270 | 2 |
| 4 | 330 | 1 |
| 5 | 390 | 3 |
| 6 | 450 | 1 |
| 7 | 500 | 3 |
Toxicity of morphine. Owing to the small number of animals surviving, I found it impossible in this preliminary experiment to establish accurately the 50 per cent lethal dose. The experiments gave the following results:
When injected subcutaneously, the animals at least tolerated 0.300 mg. per g., or twice the 50 per cent lethal dose for normal rats.
( β) With the intravenous method, the 2 per cent solution of morphine hydrochloride in physiological serum was injected in the internal saphena of a hind foot; and the mortality was as follows:
|
Doses (in δ per g.) |
Number of animals |
Mortality percentage |
| 100 | 5 | 0 |
| 150 | 4 | 20 |
| 175 | 4 | 100 |
As the dose of 0.100 mg. per g. did not cause any mortality, the surviving animals were used twenty-four hours after the general injection of morphine and increasing doses were administered to them.
Summary. Rats are able to tolerate daily increasing doses of morphine hydrochloride administered subcutaneously until these reach twice the 50 per cent lethal dose for normal animals. As rats appear to become habituated to morphine, I continued the experiments.
Second experiment. This was carried out with sixty-three rats, males and non-pregnant females, under laboratory conditions identical with those of the first experiment,12 except with respect to the concentrations of the solution; for when quantities less than 1 c.c. are injected, mortality is reduced and only amounts to 14 per cent. The following table shows the concentrations of the solutions in relation to the doses and the number of deaths per week:
|
Number of weeks |
Doses (in δ per g.) at the beginning of each week |
Concentration of morp hinehydrochloride solution (Percentages) |
Number of deaths weekly |
| 1 | 100 | 2 | 0 |
| 2 | 160 | 3 | 2 |
| 3 | 220 | 4 | 0 |
| 4 | 280 | 4 | 2 |
| 5 | 340 | 4 | 1 |
| 6 | 400 | 5 | 2 |
| 7 | 460 | 6 | 1 |
| 8 | 500 | 8 | 1 |
Determination of the 50 per cent lethal dose of morphine hydrochloride administered by subcutaneous and intravenous injection to rats habituated to morphine
Subcutaneous injection. The 4 per cent solution of morphine hydrochloride was administered to the rats of the second experiment after they had received 0.500 mg. per g. on the previous day. The table below shows the mortality percentage obtained in twenty-four hours in relation to the doses injected.13
Determination of the toxicity of morphine hydrochloride administered by subcutaneous injection to rats habituated to morphine
|
Doses (in δ per g.) |
Number of animals |
Mortality percentage |
| 600 | 5 | 0 |
| 650 | 5 | 0 |
| 700 | 5 | 0 |
| 750 | 6 | 0 |
| 800 | 10 | 50 |
These animals will be referred to as group III
13During the experiment the animals not used for the determination of toxicity continue to receive 0.500 mg. per g. daily.
Average variation for the 50 per cent lethal dose: 16. 50 per cent lethal dose of morphine hydrochloride administered by subcutaneous injection to rats habituated to morphine for seven weeks: 800 ± 10 δ per g., or 320 per cent of the dose 50 per cent lethal to normal animals. Percentage of error: 1.2 per cent.
Intravenous injection. The injection is administered in the internal saphena of a hind foot with a 2 per cent solution of morphine hydrochloride in physiological serum. The mortality percentages twenty-four hours after the injection of morphine appear in the table below.14
|
Doses (in δ per g.) |
Number of animals |
Mortality percentage |
| 100 | 5 | 0 |
| 150 | 10 | 10 |
| 160 | 5 | 20 |
| 170 | 6 | 100 |
| 175 | 4 | 100 |
Average variation for the 50 per cent lethal dose: 16.50 per cent lethal dose of morphine hydrochloride administered by intravenous injection to habituated animals: 164 ± 19 δ per g., or 164 per cent of the dose 50 per cent lethal to normal animals.
Percentage of error: 11.5 per cent.
Toxicity by subcutaneous injection/toxicity by intravenous injection. In rats this ratio amounts to 4.8. Rats are able to tolerate by subcutaneous injection a dose equivalent to more than three times the dose toxic for normal animals and by intravenous injection a dose equivalent to more than 1.5 times the dose toxic for normal animals. Furthermore, since the ratio of toxicities according to the method of injection is 4.8, morphine. seems to undergo a more rapid conversion in the organisms of rats habituated to the drug than in those of animals which have not been injected. It approaches the similar ratio obtained with oxydimorphine.
Control experiment. In order to ascertain whether tolerance of morphine is not caused by the effect of the injection alone, twenty-four rats were administered 0.33 c.c. of distilled water subcutaneously daily for three weeks. After that period, the 50 per cent dose of morphine hydrochloride injected subcutaneously was determined in the case of ten of these rats.
Mortality amounted to 50 per cent for 0.250 mg. per g., and is therefore identical with that obtained with normal animals.
This experiment confirms that tolerance of large doses of morphine is due to the effect of the repeated injections of morphine hydrochloride.
14During the experiment the animals not used for the determination of toxicity continue to receive 0.500 mg. per g. daily.
Different methods of administering the drug
I wished to determine the speed with which rats are able to tolerate toxic doses of morphine. I shall describe below the various experiments carried out on groups of animals submitted to chronic intoxication of greater or less intensity for varying lengths of time:
Group I (three days). Subcutaneous injections of 0.100 mg. of morphine hydrochloride per g. of weight are administered to thirty rats for three consecutive days. A 2 per cent concentration of the solution is used. The toxicities are determined on the fourth day. The mortality observed in relation to the doses and the methods of injection are summed up in the table below:
|
Doses |
Number of animals |
Mortality percentage |
|---|---|---|
|
Subcutaneous injection: |
|
|
| 600 | 6 | 0 |
| 800 | 6 | 18 |
|
Intravenous injection: |
|
|
| 150 | 6 | 18 |
| 160 | 5 | 20 |
| 170 | 2 | 100 |
These rats tolerate the same doses (0.800 mg. per g. with subcutaneous injection, 0.165 mg. per g. with intravenous injection) as animals habituated to morphine for seven weeks. This tolerance of large doses appeared to be surprising, but I was able to confirm this phenomenon by numerous trials.
Group II (three weeks). Very rapidly increasing doses of morphine hydrochloride were administered by subcutaneous injection to fifty-four rats daily (except Sundays). The appended table shows the successive doses injected, the concentrations of the solutions, the number of animals to which injections were administered and the mortality. A dose of 0.500 mg. per g. is reached, and is then administered daily to a number of rats, while others are given increasing doses in order to determine the maximum quantity tolerated.
The low mortality proves that rats become habituated very rapidly to morphine. After receiving injections for twenty-eight days, four rats tolerated four times the dose 50 per cent lethal to normal animals.
|
Number of weeks |
Doses in δ per g. |
Concentrations (Percentages) |
Number of animals injected |
Number of deaths |
| 1 | 100 | 2 | 54 |
|
|
|
120 | 2 | 54 |
|
|
|
140 | 2 | 54 |
|
|
|
160 | 4 | 53 | 1 |
|
|
180 | 4 | 53 |
|
|
|
200 | 4 | 53 |
|
| 2 | 220 | 4 | 53 |
|
|
|
250 | 4 | 53 |
|
|
|
300 | 6 | 53 |
|
|
|
350 | 6 | 53 |
|
|
|
400 | 8 | 53 |
|
|
|
450 | 8 | 52 | 1 |
| 3 | 500 | 8 | 51 |
|
|
|
550 | 8 | 15 |
|
|
|
600 | 8 | 15 |
|
|
|
650 | 8 | 13 |
|
|
|
700 | 8 | 13 |
|
| 4 | 750 | 8 | 8 | 1 |
|
|
800 | 8 | 12 | 1 |
|
|
850 | 8 | 11 | 1 |
|
|
900 | 8 | 5 | 1 |
|
|
1,000 | 8 | 4 |
On the basis of the results obtained with these groups I and II, it might seem unnecessary to habituate the rats for more than three weeks or even three days. This method, however, points to certain observations, as I shall show in connexion with the wearing off of the analgesic action and with elimination.
Group III (seven weeks). The method of administration and the toxicities of morphine in this group of rats has been described on pages 30 and 31. These are the animals to which injections of doses ranging from 0.100 to 0.500 mg. per g., increased each day by 0.010 mg. per g., were administered for seven weeks.
Group IV (twelve weeks). These rats are those in group III which continue to receive daily, for five weeks, 0.500 mg. of an 8 per cent morphine hydrochloride solution per g. of body weight. At the end of the tenth week the rats, with numerous scars on their flanks and rapidly diminishing weight, are dying. After twelve weeks, the animals have scars and dull shaggy fur and take very little food. The mortality rate rises. With a dose of 0.500 mg. per g., it seems difficult to keep them alive beyond this period.
Summary: The various methods of administering the morphine to the rats were as follows:
Group I: Rats receiving a subcutaneous injection of 0.100 mg. of morphine hydrochloride per g: for three consecutive days.
Group II: Rats receiving subcutaneous injections of rapidly increasing doses of morphine hydrochloride over a period of three weeks.
|
1st week: |
0.10; |
0.12; |
0.14; |
0.16; |
0.18; |
0.20; |
mg. per g. |
|
2nd week: |
0.22; |
0.25; |
0.30; |
0.35; |
0.40; |
0.45; |
mg. Per g. |
|
3rd week: |
0.50; |
0.50; |
0.50; |
0.50; |
0.50; |
0.50; |
mg. per g. |
Group III: Rats receiving subcutaneous injections of doses of morphine hydrochloride increasing by 0.01 mg. per g. daily for seven weeks.
|
Initial dose: |
0.10 mg. per g. |
|
Final dose: |
0.50 mg. per g. |
Group IV: Rats receiving subcutaneous injections of doses of morphine hydrochloride increasing by 0.01 mg. per g. daily, for twelve weeks.
|
Initial dose: |
0.10 mg. per g. |
The dose of 0.50 mg. per g. is reached seven weeks after administration of the drug begins, and the rats continue to receive this injection for five weeks.
Tolerance to toxic doses of morphine develops very quickly in rats. Three successive injections (group I) enable them to tolerate three times more morphine than the normal animal. The daily administration, for three or seven weeks (groups II and III), of increasing doses of morphine makes them capable of tolerating doses, four times greater than the normal rat. However, tolerance to large doses does not extend beyond twelve Weeks (group IV). The ratio, of toxicity according to the method of introduction (subcutaneous and intravenous), which is higher in the animal habituated to the drug than in the normal animal, might indicate a more rapid transformation of the morphine into a less toxic substance in the organism of the habituated animal.
B. Physical condition of rats habituated to morphine
This study relates to the behaviour of rats in groups II and III.
Depressive effect of morphine
In normal rats, small doses of morphine hydrochloride (0.100 mg. per g.), given by subcutaneous injection, induce drowsiness twenty minutes after the injection, and this state continues for two to three hours.
Larger doses (0.300 mg. per g.) produce, in addition to drowsiness, diminishing frequency of respiratory movement and increased depth of respiration. Death occurs in one to two hours.
In habituated rats, drowsiness after injection only appears for the first three to five days. In the second week, the rat shows no depressive symptoms after the injection, behaves like an animal which has not been injected and takes food normally.
Very large doses (0.500 mg. per g.), always induce a state of agitation in. habituated animals for one to two hours after the injection. The slightest noise makes them start.
Growth
As stated previously (page 31), growth is arrested by the regular administration of morphine hydrochloride. The following table gives, by way of example, the weights of three rats before the experiment and seven weeks later.
|
Weight before experiment |
Weight (in g.) 7 weeks later (dose injected 0.500 mg/g) |
|---|---|
| 82 | 95 |
| 84 | 82 |
| 105 | 109 |
Reproduction
The male and female genital organs of habituated rats become atrophied. The testicles and seminal vesicles lose 50 to 75 per cent in weight, and the spermatozoa disappear. In females, the uterine cornua become filiform and the ovaries have no ripe follicles. I have never noted any case of pregnancy when males and females have been put together in the same bowl.
C. Diminution of analgesic action during habituation
The test used to study this subject was that described by Mme. Buchel and Jeanne Lévy (1948-1949) This test, which was carried out on rats in order to compare the analgesic effect of various drugs, enabled me to trace, during the period of habituation, the analgesic effect of an initial active dose of morphine on the normal rat, and to determine the dose needed to induce the same state during the progress of habituation.
The analgesic power of various doses of morphine hydrochloride according to the method of introduction (subcutaneous and intraperitoneal) was first determined in normal rats.
Description of the test
The rat is tied on its back to a board by its feet and head. In this uncomfortable position it squeals and struggles. Morphine hydrochloride is administered. The length of time needed for the squealing on the one hand and the spontaneous struggling on the other to subside (shown in the tables by - +) is noted and corresponds. to a state of slight analgesia; the length of time needed for the rat to fail to react; either by squealing or struggling, to a strong pinching of the tail with Kocher's forceps (shown in the tables by -), is noted and corresponds to a state of deep analgesia. The duration represents the length of time in which the animal remains in the - state.15
Determination of the analgesic effect of morphine hydrochloride on normal rats
I give below, as an example, the calculation of the degree of analgesia produced in relation to the dose of morphine hydrochloride administered by subcutaneous and intraperitoneal injection; it was carried out on normal rats, males and non-pregnant females; aged from three to four months, and weighing 80 to 120 grammes. Each dose represents the mean effect observed in series of six animals. A 1 per cent solution of morphine hydrochloride was used.
15The duration of the + - state varies; in some experiments, it may be several hours before the rats squeal or struggle.
|
Squealing Period of latency (min) |
Agitation Period of latency (min) |
||||||
|---|---|---|---|---|---|---|---|
|
Cose injected (in δ per g.) |
- + |
- |
Duration (min) |
- + |
- |
Duration (min) | |
|
Subcutaneous |
25 | 24 | 26 | 27 | 32 | 47 | 10 |
|
|
50 | 24 | 37 | 59 | 19 | 34 | 17 |
|
|
100 | 19 | 32 | 48 | 21 | 22 | 21 |
|
|
125 | 18 | 28 | 137 | 18 | 38 | 31 |
|
|
150 | 12 | 28 | 145 | 22 | 32 | 105 |
|
Intraperitoneal |
25 | 15 | 37 | 15 | 30 | 0 | 0 |
|
|
50 | 14 | 24 | 50 | 18 | 40 | 19 |
|
|
100 | 6 | 11 | 88 | 10 | 19 | 49 |
|
|
150 | 1 | 5 | 110 | 5 | 10 | 65 |
|
|
200 | 1 | 15 | 170 | 15 | 20 | 92 |
Each calculation must be carried out on a rat which has never received analgesia, since the animal reacts less well to an active dose of morphine as soon as it has received a first injection.
The injections must be separated by a period of seven to fourteen days if the rat is to show constant behaviour towards morphine hydrochloride.
|
Squealing Period of latency (min) |
Agitation Period of latency (min) |
|||||
|---|---|---|---|---|---|---|
|
Time separating 2 injections (in days) |
- + |
- |
Duration (min) |
- + |
- |
Duration (min) |
| 0 | 3.5 | 7.5 | 108 | 3.5 | 7.5 | 50 |
| 2 | 4 | 11 | 70 | 4 | 11 | 15 |
| 0 | 3.5 | 8.5 | 137 | 3.5 | 8.5 | 49 |
| 7 | 3.5 | 9.5 | 79 | 3.5 | 9.5 | 57 |
| 0 | 3 | 10 | 96 | 3 | 10 | 45 |
| 14 | 4 | 8 | 150 | 6 | 19 | 34 |
Determination of the analgesic effect of morphine hydrochloride on rats under morphine treatment
The analgesic effect of morphine hydrochloride on rats is determined by administering the drug in steadily increasing doses, the initial dose being 0.100 mg. per g. This initial dose of 0.100 mg. per g. selected for intra- peritoneal injection induces a period of analgesia in a normal rat sufficiently prolonged to enable its effect to be determined at the outset of the period of habituation.
Analgesic effects of morphine on rats in group I.
This is measured for three days on five rats after intraperitoneal injection of 0.100 mg. per g. and on the fourth day on the same animals after intraperitoneal injection of 0.300 mg. per g. It can be seen from the following table that the period of analgesia regularly diminishes on the first three days and is still shorter on the fourth day after the injection of a dose three times greater than the original dose.
|
Squealing Period of latency (min) |
Agitation Period of latency (min) |
||||||
|---|---|---|---|---|---|---|---|
|
Number of days |
Dose injected (in δ per g.) |
- + |
- |
Duration (min) |
- + |
- |
Duration (min) |
| 1 | 100 |
2 1/3 |
9 | 94 |
2 1/3 |
9 | 60 |
| 2 | 100 |
2 1/4 |
7 1/4 |
75 |
2 1/4 |
9 | 40 |
| 3 | 100 | 4 | 8 | 31 |
5 1/2 |
7 1/2 |
13 |
| 4 | 300 | 3 | 11 | 22 | 8 | 4 | 6 |
Analgesic effect of morphine on rats in group II.
This action was determined by administering rapidly increasing doses of morphine hydrochloride by subcutaneous injection to twelve rats before habituation began. Analgesia is first measured after intraperitoneal injection of 0.100 mg. per g. into normal rats, and is then measured in the same animals after the drug has been administered for three successive days. The analgesic effect is then observed in the same rats after eleven and twenty days of injection, the doses of morphine hydrochloride administered by intraperitoneal injection at the time of measurement being 0.200, 0.300 and 0.400 mg. per g. It is clear from the following table of the average results obtained that the duration of analgesia regularly diminishes over the period of twenty days, morphine no longer having any analgesic effect at the end of that period, despite the increase in the dose of the intraperitoneal injection to four times that of the original injection.
|
Squealing Period of latency (min) |
Agitation Period of latency (min) |
||||||
|---|---|---|---|---|---|---|---|
|
Doses injected 24 hours before test (in δ/g.) |
Doses administered by intraperitoneal injection (in δ/g.) |
- + |
- |
Duration (min) |
- + |
- |
Duration (min) |
| 0 | 100 | 4 | 7 | 131 | 4 | 7 | 0 |
| 140 | 100 | 6 | 17 | 34 | 6 | 0 | 0 |
| 240 | 200 | 9 | 35 | 13 | 9 | 0 | 0 |
| 450 | 300 | 6 | 0 | 0 | 0 | 0 | 0 |
| 500 | 400 | 3 | 0 | 0 | 3 | 0 | 0 |
Analgesic effect of morphine on rats in group III.
Analgesic action is observed in four rats after intraperitoneal injection of 0.100 mg. per g. Daily increasing doses of morphine hydrochloride are then administered to these rats by subcutaneous injection, the initial dose being 0.100 mg. per g., increasing daily by 0.010 mg. per g.
The following table shows the average increase in the period of latency and the average decline in the duration of analgesia in the four rats during the period of habituation, the rats being examined each week after the intraperitoneal injection of 0.100 mg. of morphine hydrochloride per g. of body weight.
|
Squealing Period of latency (min) |
Agitation Period of latency (min) |
||||||
|---|---|---|---|---|---|---|---|
|
Doses injected 24 hours before test (in δ/g.) |
Doses administered by intraperitoneal injection (in δ/g.) |
- + |
- |
Duration (min) |
- + |
- |
Duration (min) |
|
Normal |
100 | 5 | 12 | 110 | 5 | 15 | 45 |
| 170 | 100 | 5 | 15 | 45 | 15 | 0 | 0 |
| 230 | 100 | 15 | 50 | 10 | 20 | 0 | 0 |
| 300 | 100 | 30 | 0 | 0 | 0 | 0 | 0 |
All figures represent the average results obtained by experiment.
No further analgesic effect being observed with the original dose of 0.100 mg. per g. of morphine hydrochloride, the dose of the intraperitoneal injections is increased every eight or fifteen days. It can be seen from the following table that the increasing quantities of the drug injected to measure the analgesia reduce the period of latency before the cessation of spontaneous squealing. Injections of 0.450 and 0.500 mg. per g., however, still eliminate spontaneous agitation, as well as squealing when the tail is pinched, without producing profound analgesia in highly habituated rats.
|
Squealing Period of latency (min) |
Agitation Period of latency (min) |
||||||
|---|---|---|---|---|---|---|---|
|
Doses injected 24 hours before test (in δ/g.) |
Doses administered by intraperitoneal injection (in δ/g.) |
- + |
- |
Duration (min) |
- + |
- |
Duration (min) |
| 350 | 200 | 15 | 0 | 0 | 0 | 0 | 0 |
| 400 | 200 | 15 | 0 | 0 | 0 | 0 | 0 |
| 460 | 300 | 20 | 0 | 0 | 0 | 0 | 0 |
| 520 | 300 | 10 | 0 | 0 | 0 | 0 | 0 |
| 580 | 400 | 10 | 0 | 0 | 0 | 0 | 0 |
| 620 | 450 | 5 | 20 | 15 | 15 | 0 | 0 |
| 680 | 500 | 20 | 30 | 15 | 60 | 0 | 0 |
All figures represent the average results obtained by experiment.
The effect of larger doses could not be determined because of their toxicity.
The 50 per cent lethal dose of morphine hydrochloride administered by intraperitoneal injection is 0.225 mg. per g. in normal animals and 0.600 mg. per g. in habituated animals.
Summary. The reduction and even the disappearance of the analgesic effect of morphine is similar in the case of the rats in groups II and III.
In the case of the rats in group I, the analgesic effect of morphine diminishes but has not disappeared on the fourth day; there is no parallelism between the two main tests used to define habituation -tolerance of toxic doses and reduction of the analgestic action.
In the case of the rats in groups II and III, on the other hand, the two tests to determine habituation produce comparable results.
D. De-habituation
This was studied in the rats in groups II and III for which the last injection of morphine hydrochloride was 0.500 mg. per g.
Agitation. Non-injected animals show signs of agitation on the second and third days after discontinuance of the drug and endeavour to bite if any attempt is made to catch them. On the fourth day this agitation disappears completely.
Weight. The weight of the rats diminishes for forty-eight hours after the last injection, remains unchanged on the third and fourth days and begins to increase on the fifth day. Twenty per cent of the animals,however, become very thin and die four days after discontinuance of the drug. These results are shown in the following table:
Weight of rats after discontinuance of morphine hydrochloride
|
Weight at time of last injection |
Weight during the period in which morphine is discontinued | |||
|
(in g.) |
2 days |
3 days |
4 days |
5 days |
| 109 | 100 | 100 | 98 | 103 |
| 113 | 100 | 98 | 92 | 100 |
| 121 | 102 | 100 | 100 | 105 |
| 115 | 97 | 84 |
D |
- |
| 119 | 114 | 103 | 98 | 100 |
| 103 | 98 | 79 |
D |
- |
Tolerance of morphine. The rats are able to tolerate an initial dose of 0.500 mg. per g. if the period for which the drug is withheld does not exceed five days. Beyond that period they all die without exception when such a dose is administered.
Analgesia. The analgesic effect of morphine is studied during the process of de-habituation.
A number of rats in group II which have been given a final dose of 0.500 mg. per g: of morphine hydrochloride are given an initial dose of morphine (0.100 mg. per g.) by intraperitoneal injection seven, fourteen and twenty-five days after discontinuance of morphine. Analgesic action was slight after seven days, stronger after fourteen days and almost normal after twenty-five days. The following table shows in brief· the average analgesic effect on normal rats of 0.100 mg. per g. administered by intraperitoneal injection, the analgesic effect of 0.400 mg. per g. on rats habituated for a period of three weeks, and the analgesic effect after injection of 0.100 mg. per g. into the same rats seven, fourteen and twenty-five days after discontinuance of the drug.
|
Squealing Period of latency (min) |
Agitation Period of latency (min) |
|||||||
|---|---|---|---|---|---|---|---|---|
|
Doses injected 24 hours before test (δ/g.) |
Number of days after withdrawal (δ/g.) |
Doses by intraperitoneal injection |
- + |
- |
Duration (min) |
- + |
- |
Duration (min) |
|
Normal |
|
100 | 4 | 7 | 131 | 4 | 7 | 76 |
| 500 |
|
400 | 3 | 0 | 0 | 3 | 0 | 0 |
|
|
7 | 100 | 4 | 23 | 46 | 17 | 0 | 0 |
|
|
14 | 100 | 3 | 10 | 72 | 3 | 14 | 15 |
|
|
25 | 100 | 6 | 13 | 110 | 7 | 22 | 34 |
Fourteen days after the withdrawal of the morphine, the analgesic effect is still weak and it takes at least twenty-five days for the effect to approach normal. Habituated rats behave differently from normal rats, in which the original analgesic effect reappears between seven and fourteen days after the injection.
E. Reaction to hypnotics of rats habituated to morphine
This study was carried out in order to show that a strongly habituated animal may fail to show cross habituation to hypnotics.
Research into the wearing off of the effect of hypnotics on animals habituated to morphine has led scientists to contradictory results. Biberfeld (1916) with veronal, Myers (1916) using chloral, and Joel and Ettinger (1926) using chloral, amylene hydrate and gardenal, didnot observe clear habituation to these drugs in animals habituated to morphine. On the other hand, Tatum, Collins and Seevers (1929) found that the effect of barbiturates was less on habituated animals than on normal animals. L. Buchel having recorded modifications in the duration of sleep induced in normal rats by evipan, after previous injection of morphine, I resumed in collaboration with her this study of habituated rats.
I chose two barbiturates, evipan (N-methyl-methyl-cyclo-hexenyl-5-5-barbituric acid) and soneryl (butyl-ethyl-barbituric acid), the first inducing, in normal rats, a short and immediate period of sleep and the other a longer period of sleep induced after a certain period of latency.
Behaviour of normal rats under the influence of evipan
The three-months-old rats receive a subcutaneous injection of morphine hydrochloride, dose 0.050 or 0.200 mg. per g., or two successive injections of morphine hydrochloride at an interval of fourhours, doses 0.035 mg., 0.050 mg., 0.075 mg. or 0.100 mg. per g. Four hours and forty-eight hours after the last dose of morphine hydrochloride, they are given an intravenous injection of sodium evipan, dose 0.025 mg. per g. The average period of sleep induced by the same dose of evipan in rats of the same age which have not received a previous injection of morphine is six minutes.
|
Average duration of sleep induced by evipan | ||||
|---|---|---|---|---|
|
Number of injections |
Morphine doses (δ/g.) |
4 hours after morphine (min.) |
48 hours after morphine (min.) |
Control (min.) |
|
|
|
|
|
6 |
| 1 | 50 | 6? | 6? |
|
|
|
200 | 15 | 6? |
|
| 2 | 35 | 9? | 6 |
|
|
|
50 | 10 | 6 |
|
|
|
75 | 10? | 10? |
|
|
|
100 | 24 | 8? |
|
*The experiments were carried out on groups of five rats, and the figures represent the. average result obtained.
A prolongation of the period of sleep induced by evipan is observed in rats receiving two successive doses of morphine hydrochloride at an interval of four hours. The largest doses, 0.075 and 0.100 mg. per g., induce a slight prolongation of the period of sleep due to evipan forty-eight hours after the morphine.
If the evipan is injected after a single dose of morphine, the prolongation of the period of sleep after the largest dose of morphine (0.200 mg. per g.) is less than in the case of rats receiving two injections of 0.100 mg. per g. of morphine hydrochloride.
Behaviour of habituated animals under the influence of evipan
The same experiment is carried out on habituated rats. These are given increasing doses of morphine hydrochloride for seven weeks, according to the experimental procedure described for group III, the last dose injected being 0.500 mg. per g. The injections of sodium evipan (0.025 mg. per g.) are given four, twenty-four and forty-eight hours after the last injection of morphine hydrochloride. At the same time,normal rats of the same age are given the same dose of evipan, which produces an average period of sleep of six minutes. The average period of sleep induced by evipan in the habituated rats is greater as compared with control rats when the evipan is injected four hours after the morphine. The increase in the period of sleep is smaller when the evipan is injected twenty-four hours after the morphine. The period of sleep induced by evipan is identical with that of the control animals when the evipan is injected forty-eight hours after the morphine.
The following table gives these results:
|
Morphine (δ per g.) in habituated rats: last injection |
Time lapse between morphine injection and evipan (h.) - |
Average duration of sleep in habituated rats(min. ) |
Average duration of sleep in controls (min.) |
|
|
|
|
6 |
| 500 | 4 | 13 |
|
| 500 | 24 | 11 |
|
| 500 | 48 | 6 |
|
Experiments carried out on groups of five rats. The figures represent the average result obtained.
The average period of sleep induced by evipan is slightly less in habituated rats four hours after the administration of 0.500 mg. of morphine hydrochloride per g. than in normal rats four hours after a single injection of 0.200 mg. per g. or two successive injections of 0.100 mg. per g. Forty-eight hours after the morphine, the average period of sleep induced by evipan is identical in the control animals and in the habituated rats, while it is slightly longer in normal animals which have been given an injection of 0.200 mg. per g. or two injections of 0.075 and 0.100 mg. per g.
There is no cross habituation.
Behaviour of normal rats under the influence of soneryl
Three-months-old rats are given a subcutaneous injection of morphine hydrochloride, dose 0.200 mg. per g., or two successive injections of morphine hydrochloride at an interval of four hours, doses 0.035 mg., 0.050 mg., 0.075 mg. and 0.100 mg. per g. They are given an intravenous injection of 0.050 mg. per g. of soneryl, four hours and forty-eight hours after the last dose of morphine. The average duration of sleep16induced by the same dose of soneryl in rats of the same age which have not received a previous injection of morphine is twenty-eight minutes; individual variations are quite large, oscillating between thirteen and forty-one minutes. Sleep is not immediate and the average period of latency is five minutes; the limits of the period of latency are two to twenty-one minutes.
|
Period of latency (min.) |
Duration of sleep (min.) |
Controls | |||||
|---|---|---|---|---|---|---|---|
|
Morphine, number of injections |
Doses δ/g. |
4 hours after morphine |
48 hours after morphine |
4 hours after morphine |
48 hours after morphine |
Period of latency (min.) |
Duration of sleep (min.) |
| 5 | 28 | ||||||
| 1 | 200 | 0 | 3 | 117 | 27 | ||
| 2 | 35 |
2 1/3 |
2 3/4 |
39 1/2 |
40 3/4 |
||
| 50 |
1/2 |
6 | 106 | 36 | |||
| 75 |
1/4 |
4 1/4 |
98 | 44 | |||
| 100 | 0 |
4 1/2 |
74 | 27 | |||
*The experiments were carried out on groups of five rats.
**The experiments were carried out on twenty-eight rats.
An injection of 0.200 mg. per g. of morphine hydrochloride does not alter the length of sleep caused by soneryl. After the injection of two doses of 0.050, 0.075 and 0.100 mg. per g., the period of sleep caused by soneryl is increased if the soneryl is injected four hours after the morphine, but the period of sleep caused by soneryl injected forty-eight hours after the morphine is similar to that observed in the control rats.
Behaviour of habituated rats under the influence of soneryl
The same experiment is carried out with habituated rats which have received constant doses of morphine hydrochloride varying from 0.100 mg. to 0.500 mg. for three weeks (rats in group II). Doses of 0.050 mg. ofsoneryl are administered intravenously four, twenty-four and forty-eight hours after 0.500 mg. of morphine hydrochloride per g. The average period of sleep caused by soneryl is much increased when the injection is made four hours after the morphine, but it is similar to that of the control rats twenty-four and forty-eight hours after the morphine.
The average length of sleep caused by soneryl is approximately the same in habituated rats having received 0.500 mg. of morphine hydrochloride per g. and in normal rats having received a single injection of morphine at 0.200 mg. per g. of morphine or two injections of 0.050 and 0.075 mg. per g. There again there is here no cross habituation.
|
Controls | |||||
|---|---|---|---|---|---|
|
Morphine, last injections (in δ/g) |
Lapse of time between injection of morphine and soneryl(h) |
Period of latency (min.) |
Duration of sleep (min.) |
Period of latency (min.) |
Duration of sleep (min.) |
|
|
5 | 28 | |||
| 500 | 4 |
1/3 |
115 | ||
| 500 | 24 | 4 | 30 | ||
| 500 | 48 |
4 3/4 |
18 1/2 |
||
*Experiments were carried out on groups of five rats.
**Experiments were carried out on twenty-eight rats.
Summary. The reaction of habituated rats to the two hypnotics studied is not very different from that of normal rats having received one or two preliminary injections of morphine. In those two groups of animals the average duration of sleep is prolonged, and it cannot therefore be supposed that cross habituation has been established.
F. Habituation to heroin
I studied habituation to heroin by determining, as in the case of morphine, the tolerance of toxic doses in the normal animal and in the animal having received repeated injections of heroin.
16The experiments were carried out on twenty-eight rats.
In a second series, I examined the analgesic effects of heroin and the behaviour of rats habituated to heroin under the influence of evipan.
Determination of the 50 per cent lethal dose of diacetylmorphine hydrochloride administered by subcutaneous and intravenous injection to normal rats
Subcutaneous injection. Male and non-pregnant female animals aged three months are injected in the flank with a 2 per cent solution of diacetylmorphine hydrochloride (heroin). The results are as follows:
|
Doses in i per g. |
Number of animals |
Mortality percentage |
| 50 | 4 | 0 |
| 60 | 6 | 0 |
| 70 | 6 | 50 |
| 125 | 6 | 80 |
| 150 | 6 | 100 |
Average variation: 20.
50 per cent lethal dose by subcutaneous injection: 70± 25 ?per g.
Percentage of error: 35 per cent.
Intravenous injection. A 4 per cent solution of diacetylmorphine hydrochloride in physiological serum, is injected into the internal saphena of a hind foot. The mortality percentages for the various doses are shown in the following table:
|
Dose in i per g. |
Number of animals |
Mortality percentage |
| 20 | 5 | 20 |
| 25 | 8 | 25 |
| 27 | 6 | 33 |
| 30 | 15 | 75 |
Average variation: 17.
50 per cent lethal dose of diacetylmorphine hydrochloride administered by intravenous injections: 28 ± 4.3 per g.
Percentage of error: 15 percent.
Toxicity by subcutaneous injection/toxicity by intravenous injection. This ratio for diacetylmorphine hydrochloride amounts to 2.5 and appears to indicate that heroin like morphine only undergoes slow changes in the organism.
Daily administration of heroin to rats
Diacetylmorphine hydrochloride was administered to rats: (a) in constant small doses (0.010 mg. per g.); (b) in increasing doses.
Administration of constant doses of heroin. Twelve rats, males and non-pregnant females, aged three months, received daily subcutaneous injections of a dose not toxic for the normal animal. The injections were given over a period of nineteen days. The analgesic effect was determined every week as I shall show later. The concentration of the solution was 2 per cent and the
dose injected was 0.010 mg. per g. Two of the rats died after the third dose and six out of ten rats after the nineteenth dose. The animals were in a drowsy state after each injection. Subcutaneous injection of 0.125 mg. per g. of diacetylmorphine hydrochloride and intravenous injections of 0.030 mg. per g. did not cause any deaths among the four surviving rats. Despite their low resistance to repeated injections of heroin, they withstood larger doses than those which would be toxic for normal animals,.whether administered by subcutaneous or intravenous injection.
Administration of increasing doses of heroin. Twenty-four rats, male and female, were given daily subcutaneous injections of increasing doses of diacetylmorphine hydrochloride. The initial dose was 0.010mg. per g.; on the thirtieth day they received 0.070 mg. per g. The doses were increased daily by 0.002 mg. per g. After thirty days the injections were continued on a few rats which tolerated up to 0.100 mg. per g.; successive doses of 0.070, 0.075, 0.080, 0.090 and 0.100 mg. per g. were injected. During that final period, no deaths were observed.
The concentration of the solutions in relation to the doses and the mortality during the period of habituation of the rats are shown in the table below, together with the number of animals injected.
|
Doses of heroin injected i per g.) |
Concentration of solution (percentage |
Number of animals injected |
Number of deaths |
| 10 | 2 | 24 |
- |
| 16 | 2 | 24 | 1 |
| 18 | 4 | 23 | 1 |
| 28 | 6 | 22 | 2 |
| 34 | 8 | 20 |
- |
| 46 | 1 | 20 | 1 |
| 70 | 1 | 19 | 1 |
| 100 | 1 | 6 | 0 |
The animals behaved in exactly the same way as those to which morphine had been administered: they were drowsy the first four days after the injection, and in a normal state thereafter.
The growth of the animals is arrested by the daily administration of heroin.
Determination of the 50 per cent lethal dose administered by subcutaneous and intravenous injection
Subcutaneous injection. A 2 per cent solution of diacetylmorphine hydrochloride was injected subcutaneously in rats which had been given a last dose of 0.070 mg. per g. during the period of habituation. In view of the small number of animals surviving, they had to be given in succession the different doses of diacetylmorphine hydrochloride.
|
Dose (i per g.) |
Number of animals |
Mortality percentage |
| 200 | 6 | 0 |
| 250 | 4 | 0 |
| 300 | 5 | 0 |
| 350 | 14 | 14 |
| 400 | 5 | 20 |
| 450 | 4 | 0 |
| 500 | 4 | 0 |
| 600 | 8 | 0 |
| 700 | 4 | 75 |
Average variation: 17.
50 per cent lethal dos e of diacetylmorphine hydrochloride administered by subcutaneous injection: 666 ± 200 per g.
Percentage of error: 30 per cent.
Intravenous injection. Injection of a 1 per cent solution of diacetylmorphine hydrochloride in a physiological serum in the internal saphena of a hind foot gave the following mortality percentages according to the dose:
|
Dose (i per g.) |
Number of animals |
Mortality percentage |
| 30 | 5 | 20 |
| 40 | 5 | 20 |
| 45 | 8 | 25 |
| 50 | 5 | 100 |
Average variation: 17.
50 per cent lethal dose of diacetylmorphine hydrochloride administered by intravenous injection: 466 ± 66 units δ per g.
Percentage of error: 14 percent.
Toxicity by subcutaneous injection/toxicity by intravenous injection. This ratio for diacetylmorphine hydrochloride in rats habituated to the drug amounts to 14.2. Compared to the figure for normal animals, this high value seems to indicate a more·rapid transformationof heroin in the organism of the rat habituated to the drug than in the normal animal.
Since the 50 per cent lethal doses by subcutaneous injection of heroin and morphine are almost the same for animals habituated to the drug, it seems likely that heroin undergoes an intermediary transformation into morphine through disacetylation. Wright (1940) observed, moreover, that heroin mixed with human serum is slowly disacetylated.
Summary. Rats can only tolerate a small but repeated dose of heroin for a few days. After nineteen daily injections of 0.010 mg. per g., however, they can tolerate a subcutaneous injection of a dose double that which would be toxic for the normal animal. Animals receiving increasing daily doses of heroin can tolerate a subcutaneous injection of a quantity of heroin ten times greater than that which would be toxic for the normal animal.
It maybe said that from the point of·view of.toxicity17 rats become habituated to heroin. They have a greater toleration of heroin than of morphine: they withstand subcutaneous injections of ten times more heroin and only four times more morphine than the normal rat. In man, toleration of heroin is also greater than toleration of morphine (Biggam, Arafa and Ragab, 1932).
Diminution of the analgesic effect during habituation
The analgesic effect of heroin has been studied by means of the same test as that applied to morphine by L. Buchel and Jeanne Lévy (1949). Heroin is much more active than morphine, particularly the initial dose which, administered by interperitoneal injection is 0.007 mg. per g. for heroin and 0.025 mg. per g. for morphine.
Analgesic effect of heroin on the normal rat. The table below shows the results obtained with this drug according to the method of injection.
The figures represent an average of at least four experiments
|
Squealing* Period of latency (min.) |
Agitation* Period of latency (min.) |
|||||
|---|---|---|---|---|---|---|
|
Dose injected (δ/g) |
-+ |
- |
Duration (min.) |
-+ |
- |
Duration (min.) |
| Subcutaneous |
|
|
|
|
|
|
| 1.5 | 7 |
- |
- |
9 |
- |
- |
| 3.5 | 7 | 17 | 33 | 7 | 18 | 15 |
| 7 | 9 | 15 | 33 | 7 | 21 | 30 |
| 12.5 | 7 | 12 | 50 | 7 | 12 | 55 |
| 25 | 1 | 5 | 80 | 5 | 10 | 75 |
| 50 | 1 | 5 | 126 | 4 | 9 | 122 |
|
Intraperitoneal |
|
|
|
|
|
|
| 5 | 6 |
-- |
- |
5 |
- |
- |
| 7 | 6 | 9 | 38 | 6 | 9 | 22 |
| 10 | 1 | 5 | 30 | 1 | 9 | 18 |
| 25 | 1 | 4 | 55 | 1 | 5 | 14 |
| 50 | 1 | 6 | 70 | 1 | 5 | 57 |
| 100 | 1 | 5 | 135 | 1 | 5 | 60 |
*The figures represent an average of at least four experiments.
The analgesic effect of heroin is more marked when the drug is injected subcutaneously. I carried out experiments on habituated rats with that type of injection. As the animal reacts less well after a second injection of heroin, in order to obtain equivalent analgesic·reactions, injections should be given at least nine days apart.
Analgesic effect of heroin on rats receiving regular doses of the drug. The analgesic effect was determined before the beginning of injections and twice during the period of habituation in rats injected with a constant dose of 0.010 mg. per g. of heroin and those receiving increasing doses.
A dose of 0.007 mg. per g.,18 injected subcutaneously and having an analgesic effect of rather long duration, was administered before the beginning of habituation.
The analgesic effect was observed·for the same drug after eight days of daily injections of the drug and the effect of a stronger dose (0.010 mg. per g.)19 after a second period of eight days. The tables below show the analgesic effect on the groups of four rats examined.
181 per cent solution.
192 per cent solution.
|
Squealing* Period of latency (min.) |
Agitation* Period of latency (min.) |
||||||
|---|---|---|---|---|---|---|---|
|
Number of day during which heroin was administered |
Dose injected(δ/g) subcutaneously |
-+ |
- |
Duration (min.) |
-+ |
- |
Duration (min.) |
| 0 | 7 | 7 | 12 | 124 | 10 | 17 | 81 |
| 7 | 7 | 7 | 10 | 60 | 8 | 25 | 20 |
| 14 | 10 | 4 | 9 | 50 | 7 | 15 | 40 |
*The figures represent the average of the experiment.
|
Squealing* Period of latency (min.) |
Agitation* Period of latency (min.) |
||||||
|---|---|---|---|---|---|---|---|
|
Number of day during which heroin was administered |
Dose injected(δ/g) subcutaneously |
-+ |
- |
Duration (min.) |
-+ |
- |
Duration (min.) |
| 0 | 7 | 4.5 | 9 | 115 | 5 | 13 | 66 |
| 25 | 7 | 5 | 11 | 24 | 20 |
- |
- |
| 55 | 10 | 5 | 9 | 30 | 4 | 11 | 9 |
*The figures given represent an average for all experiments.
The duration of deep analgesia diminishes even with a larger dose (0.010 mg. per g.) after fourteen days of regular administration.
The same dose of heroin administered to the animal before and after the injection of increasing doses for eight days causes a reduction in the duration of the analgesia after that period or eliminates the analgesic effect altogether, and increases the period of latency. A larger dose (0.010 mg. per g.) after fourteen days of regular, administration of increasing doses still has some effect, but it is Weak.
Summary. The analgesic effect diminishes during the habituation of rats to heroin. The diminution is greater in rats injected with increasing doses of the drug than in those to which a constant small dose is administered.
Behaviour of rats habituated to heroin under the influence of evipan
L. Buchel studied the changes in the duration of sleep caused by evipan in normal rats having previously received two injections of heroin; we made the same study with rats habituated to heroin.
Behaviour of normal rats under the influence of evipan. Injections of heroin are made subcutaneously
and evipan injections are given intravenously four hours, and forty-eight hours respectively after the second heroin injection. The average duration of sleep caused in animals of the same age by evipan alone is five minutes.
|
Heroin |
4 hours after heroin |
48 hours after heroin |
|
|
Dose (δ per g.) |
(min.) |
(min.) |
Controls |
|
|
|
|
5 |
| 4 | 6 | 5 |
|
| 7 | 17 | 12 |
|
The experiments were performed on groups of five rats and the figures show the average of the results obtained.
The above table shows that small doses of heroin (0.004 mg. per g.) do not affect the average duration of sleep induced by evipan in rats. With a large dose, the increase in the average length of sleep persists when evipan is injected forty-eight hours after two doses of heroin.
Reaction to evipan of rats habituated to heroin. Animals injected with increasing doses20 of heroin receive intravenous injections of evipan sodium four, twenty-four and forty-eight hours after the last heroin injection. Rats of the same age are injected with evipan alone.
The doses increased from 0.01 to 0.1 mg. per g., a daily increase of 0.01 mg. per g.
|
Average duration of sleep in habituated rats | |||
|---|---|---|---|
|
Heroin (δ per g.) last dose before evipan |
Time elapsed between injections of heroin and evipan (hours) |
(min.) |
Controls |
|
|
|
|
5 |
| 70 | 5 | 12 |
|
| 70 | 24 | 7 |
|
| 70 | 48 |
81/2 |
|
*The experiments were performed on groups of·five rats; figures are average·results.
The increase in the average duration of sleep is smaller if the evipan is injected·twenty-four hours after heroin than if it is administered after four hours.
Summary. A comparison of the behaviour of normal rats and those habituated to heroin under the influence of evipan shows that the increase in the length of sleep continues for a shorter period in the case of the latter than in the case of the former, despite the injection of a dose of heroin five times greater in the habituated animals than in normal rats. There is, however, no cross habituation.
G. Summary
The systematic study of the toleration by rats habituated to morphine of doses toxic to normal rats and of the wearing off of the analgesic effect of the drug indicates that the rat becomes habituated to increasing doses of morphine hydrochloride (0.100 to 0.500 mg. per g.) injected subcutaneously from three to seven weeks (groups II and III).
The 50 per cent lethal doses of morphine hydrochloride injected subcutaneously and intravenously are as follows:
250 and 0.100 mg. per g. for the normal rat;
800 and 0.165 mg. per g. for the habituated rat.
The ratio of toxicities of morphine hydrochloride, depending on how it is administered, is higher for the habituated rat (4.8) than for the normal rat (2.5) and is close to that obtained with oxydimorphine hydrochloride injected into the normal rat (6.3). Those facts consequently do not invalidate the hypothesis that morphine is converted into oxydimorphine in the habituated animal.
Toleration of toxic doses injected subcutaneously may even be built up very rapidly: after the injection of 0.100 mg. per g. for three consecutive days (group I), the rat withstands a dose three times that of the 50 per cent lethal dose injected subcutaneously for the normal animal. Those rats have the same toleration as rats which have had daily injections of increasing doses for three or seven weeks. Animals injected with increasing doses can withstand for four to five weeks daily doses 100 per cent (0.500 mg. per g.) greater than the 50 per cent lethal dose administered by subcutaneous injection (0.250 mg. per g.) for normal rats.
The analgesic effect of morphine hydrochloride injected intraperitoneally has fallen by 60 per cent eight days after the beginning of daily injections of increasing doses of morphine. At the end of three weeks, it has no effect at all and analgesic action only reappears, and then for a short period only, when a dose is administered five times as strong as that injected intraperitoneally in normal rats (groups II and III).
The weakening of the analgesic effect of morphine is noticeable on the fourth day for a dose three times as strong as the initial dose for rats in group I.
Habituated rats react to hypnotics in very much the same way as normal rats which have had two successive injections of morphine hydrochloride; there is no cross habituation.
Rats injected subcutaneously with increasing doses of heroin (0.010 to 0.100. mg. per g.), withstand a dose ten times stronger than normal animals; the high degree of toxicity, depending on how the drug is administered (subcutaneously or intravenously) in those animals seems to indicate a rapid transformation of the heroin into a less toxic substance, morphine, in the body of animals habituated to the drug. The analgesic effect of an average dose of heroin diminishes after eight days of daily injections. The rat therefore becomes habituated to regular injections of increasing doses of heroin. There is no cross habituation to the action of evipan. Rats tolerate less well repeated injections of regular weak doses of heroin (0.010 mg. per g.). Nevertheless they can tolerate without mortality a subcutaneous injection of a dose 100 per cent stronger than that which would be toxic for the normal animal, and the analgesic effect of heroin becomes weaker in those animals.
If habituation is defined in terms of the normal animal's tolerance of toxic doses and of the disappearance of the analgesic effect of a dose which was originally effective and its reappearance for a larger dose, it may be concluded that the rat is capable of becoming habituated to both morphine and heroin.
All of this work required habituating more than 600 rats.

