ABSTRACT
Introduction
Material and methods
Results
Discussion
Author: J. C. TURNER , J. K. HEMPHILL , P. G. MAHLBERG
Pages: 59 to 69
Creation Date: 1981/01/01
Gland number and cannabinoid content for three clones of Cannabiswere compared. Cannabinoid levels and mature glands were quantified throughout progressive stages of pistillate bract development in order to define possible interrelationships between glands and cannabinoids. Similar trends occurred among the clones for each gland type while individual cannabinoids displayed a different pattern in each clone. Gland initiation and development as well as cannabinoid synthesis were found to occur during bract ontogeny for all three clones. A positive correlation existed for the total number of capitate glands per bract compared to the total cannabinoid content of the bract. The study also suggested that the glands may contain the majority of the cannabinoids present in the bract.
The variability of the cannabinoid content, both qualitatively and quantitatively, found among the numerous strains of Cannabisis generally acknowledged. Aside from differences: induced by sampling techniques (Fairbairn and Liebmann, 1974; Turner, Hemphill and Mahlberg, 1977), factors controlling the cannabinoid profile in the plant are only partially understood, with both genetic and environmental influences implicated (Doorenbos et al., 1971; Bazzaz et al., 1975). Although stress has been shown to increase levels of Δ 9-tetrahydrocannabinol (Haney and Kutscheid, 1973; Coffman and Gentner, 1975), a more general theory is that the qualitative aspects are under genetic control and that the quantitative aspects are influenced by environmental factors (Fairbairn and Liebmann, 1974; Latta and Eaton, 1975).
* This research was supported by research grants from the Department of Agriculture of the United States of America and the National Institute on Drug Abuse to P. G. Mahlberg.
Several studies of Cannabis have suggested that cannabinoids are accumulated primarily in the epidermal glandular trichomes (Fujita et al., 1967; Fairbairn, 1972; Hammond and Mahlberg, 1973; DePasquale, 1974; Ledbetter and Krikorian, 1975; Malingre et al., 1975; Sharma, t975; André and Vercruysse, 1976; Small and Cronquist, 1976). Ongoing research in our laboratory has been directed toward the role of these glands as related to the cannabinoid profile of the plant (Turner et al., 1977, Turner, Hemphill and Mahlberg, 1978). In our initial study, a simple correlation between gland density and cannabinoid concentration was not apparent, but the glandular system was determined to be more complex than originally expected. Pistillate bracts were found to contain glands of varying ages (Turner et al., 1977). Also cannabinoid content in the individual glands was found to vary with gland age, gland type and location of the gland on the plant (Turner et al., 1977, 1978), In the current study, cannabinoid levels and mature glands were quantified throughout progressive stages of bract development in order to define any possible interrelationships between glands and cannabinoids.
Plants of three Cannabis strains were selected and cloned (Turner el al., 1977). These strains were derived from a high Δ 9-tetrahydrocannabinol (Δ 9- THC) strain (152), a low Δ 9-THC, high cannabidiol (CBD) strain (79), and a high CBD strain (87), Clones were grown under ambient greenhouse conditions.
Maturing bracts from pistillate plants of each clone were collected for analysis. Bracts sampled were selected at 1 mm intervals from 2 mm (very young) to 9 mm (mature) in length. A minimum of three bracts of each length was processed for scanning electron microscopy (SEM) while many bracts of each length were processed for analysis by gas-liquid chromatography (GLC). Bract samples were collected at 1500 h on three consecutive days (one clone per day) during late October 1979.
Plant parts to be analysed were processed as described previously (Turner et al., 1977). Analyses by GLC were performed on a Hewlett-Packard 5710A chromatograph equipped with a 3380A H-P integrator. Samples prepared for SEM were examined with an ETEC Autoscan.
Gland number per unit area was determined by counting glands directly on the SEM screen (Turner et al., 1977). One-half of the total surface area of each bract, from tip to base, was quantified. Measurements of surface area encompassed a minimum of 2 mm 2(32 fields) for the 2 mm bracts and a maximum of 42 mm 2(672 fields) for the 9 mm bracts. The data for each bract length sample represent a mean, and were calculated as glands per mm 2and also as total glands per bract. A minimum of three bracts for each sample length was used for the gland counts.
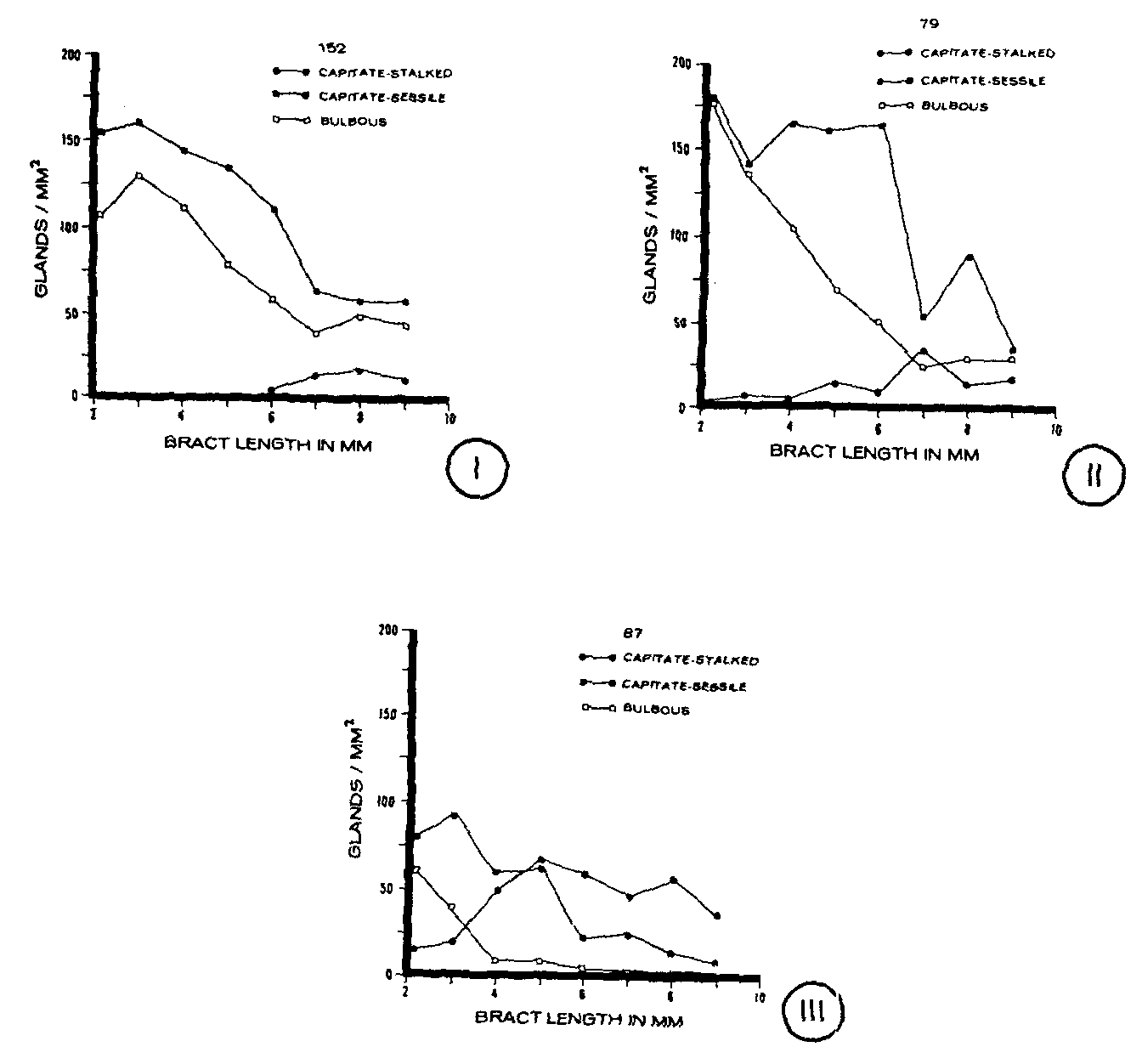
A comparison of gland densities during bract ontogeny indicated that similar trends occurred among the clones for each gland type (figure I-figure III). For all three clones, the density of capitate-stalked glands increased as the bract developed. However, the increase occurred at a different developmental stage for each clone. Capitate-stalked glands were absent from young bracts of clone 152 and appeared only after the bracts had attained a length of 6 mm (figure I). In contrast, for clones 79 and 87, capitate-stalked glands were present on the youngest bracts sampled (figure II,figure III). On clone 87, the density of this gland type increased during the early developmental stages whereas for clone 79 gland density increased during later stages of bract development. There also were numerical differences among the clones when the densities of capitate-stalked glands were compared. Both clones 152 and 79 had relatively low gland densities (figure I,figure II), while clone 87 was found to have the highest density of capitate-stalked glands (figure III).
The density of capitate-sessile glands on both clones 152 and 87 decreased as the bracts increased in length from 2 mm to 6 mm (figures I, III). The number of these glands per unit area then remained relatively constant as the bracts matured to 9 mm in length. In contrast, the density of capitate-sessile glands on clone 79 remained relatively constant until the bracts had developed to about 6 mm in length whereupon gland density decreased with further bracteal development (figure II). Quantitatively, clones 152 and 79 had similar densities of capitate-sessile glands while clone 87 had a lower gland density (figure I-figure III).
The density of bulbous glands decreased on each of the clones during early to middle stages of bract ontogeny and then remained at a relatively constant number per mm 2as the bracts approached maturity (figure I-figure III). However, while the number of bulbous glands per mm 2on clones 152 and 79 decreased gradually until the bracts were about 7 mm in length (figure I,figure II, the gland density of this gland type on clone 87 decreased only during the earliest stages of bract development (figure III). Also, clone 87 was found to have a lower density of bulbous glands than clones 152 and 79 (figures I -III).
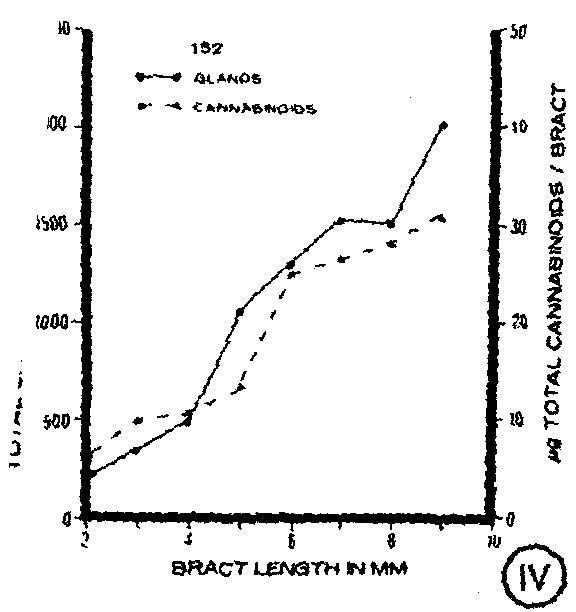
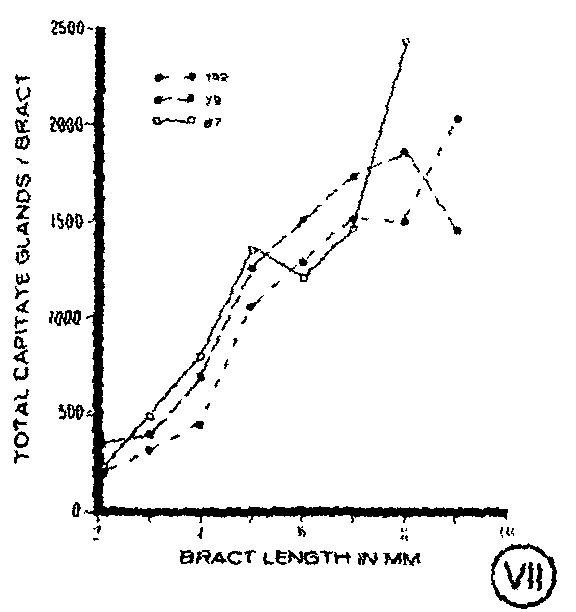
In addition to monitoring gland density during bract ontogeny, the total number of both types of capitate glands per bract was compared among the clones (figure IV-figure VII). All three clones were found to have an increase in gland number as the bract developed to maturity. When compared, the clones showed little difference in the number of capitate glands present throughout bract development until the most mature stages when quantitative differences became apparent (figure VII).
Cannabinoid concentration of developing bracts varied considerably for each of the clones (figures VIII-X). Clone 152, a drug strain which is characteristically high in Δ 9-THC, was found to have relatively similar concentrations of cannabinoids at each stage of bract development (figure VIII). The major cannabinoid detected was Δ 9-THC, averaging approximately 4 to 5 mg/100 mg dry weight, whereas cannabichromene (CBC) was detected in very small amounts. Clone 79, a non-drug strain, accumulated characteristically high levels of CBD, although a small quantity of Δ 9-THC was present as well. Clone 79 was found to have relatively similar concentrations of cannabinoids at immature stages of bract development, but as the bract matured cannabinoids increased in concentration (figure IX). Concentrations of CBD in clone 79 averaged 6 to 7 mg/100 mg dry weight or higher. In clone 87, a fibre strain which is characteristically high in CBD, cannabinoid concentration increased at each stage of bract development except for the most mature stage (figure X). As in clone 79, high levels of CBD were detected along with small concentrations of Δ 9-THC.
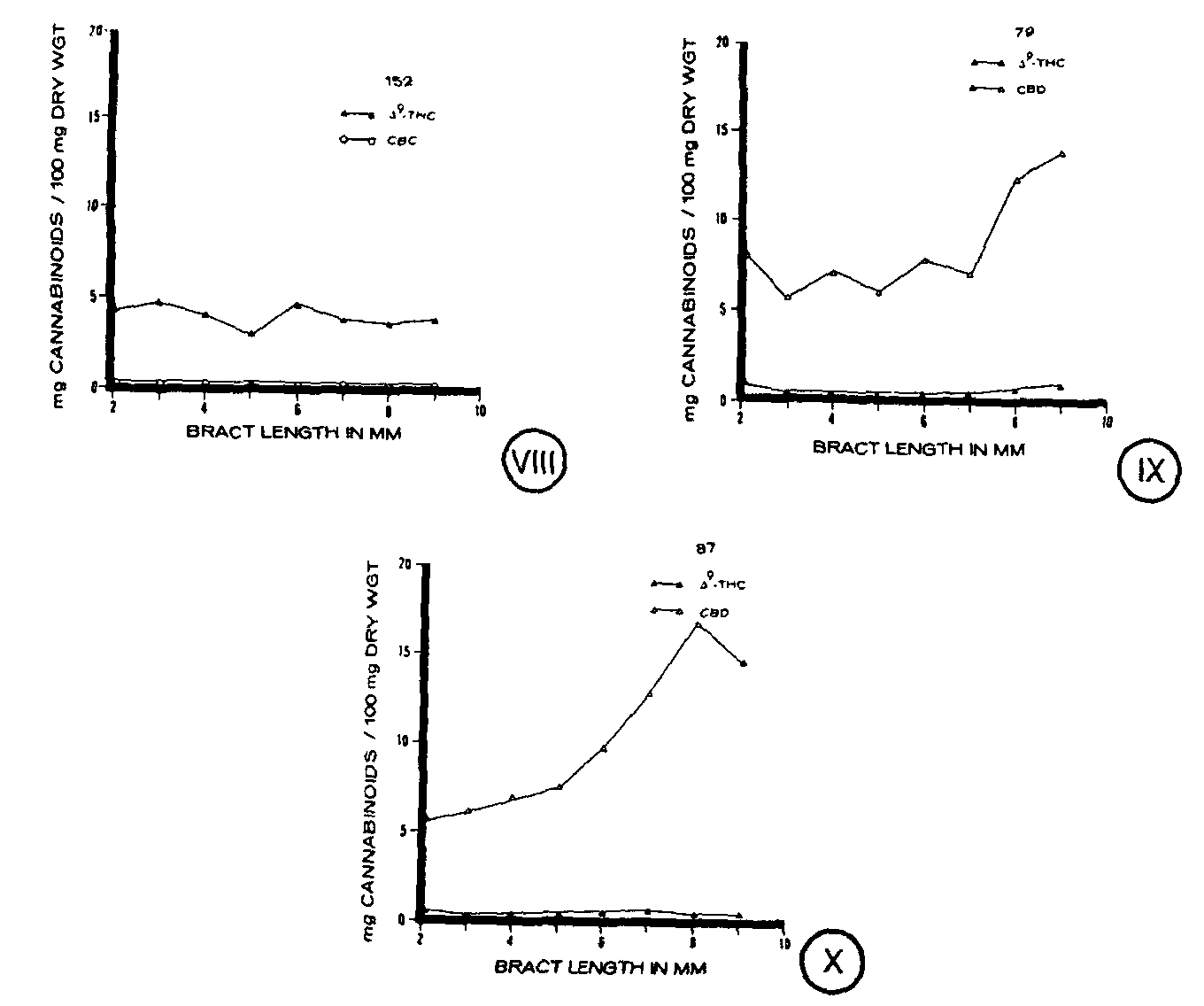
Quantities of cannabinoids detected in clone 87 were comparable with the levels found in clone 79 at all stages of bracteal development (figures IX, X).
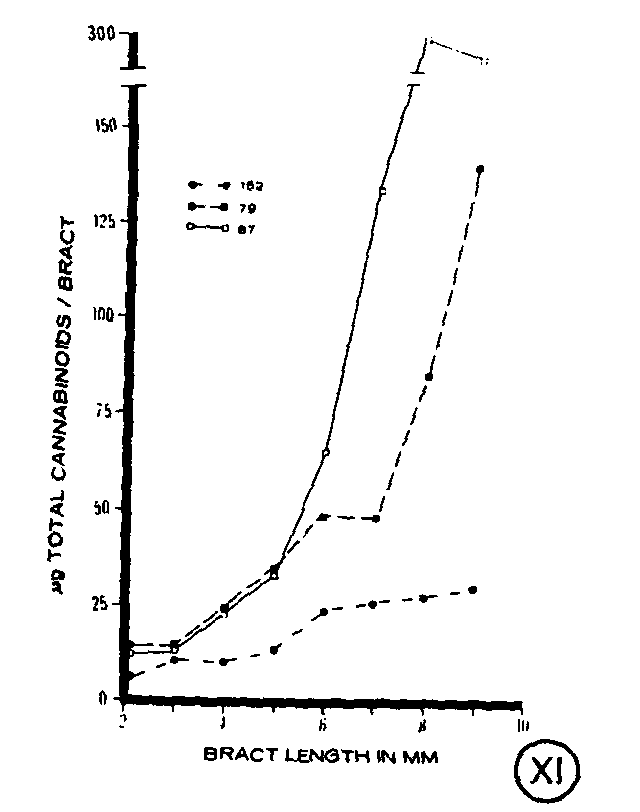
Total cannabinoid content per bract also was compared among the clones at each stage of bract development (figure XI). Differences were apparent with clone 152 having the lowest total cannabinoid content, particularly at the more mature stages. Clones 79 and 87 had similar amounts of total cannabinoids per bract present at young developmental stages, but clone 87 had considerably higher amounts of total cannabinoids per bract at the more mature stages (figure XI). A comparison of total cannabin- oid content per bract with total capitate glands per bract showed an increase m both parameters throughout bract development for each of the clones (figures IV-VI).
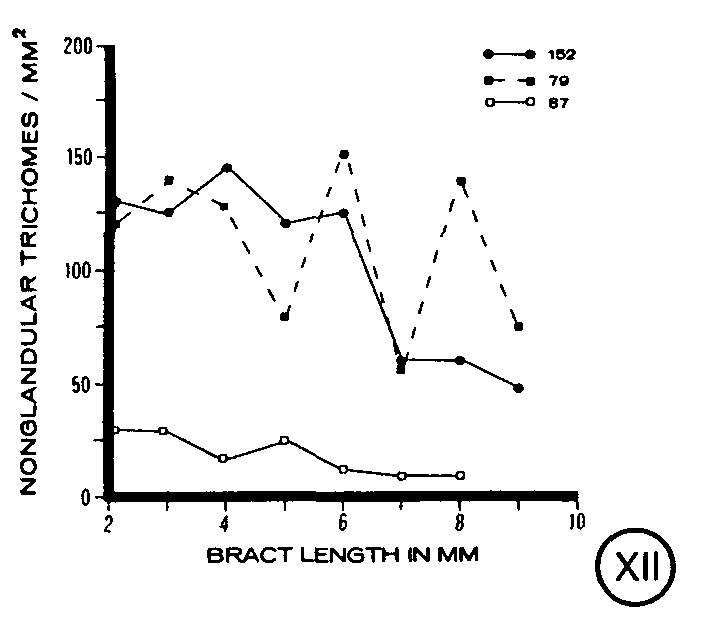
Non-glandular trichomes on clone 152 were found to maintain a uniform level of trichome density until the bract was 6 mm in length, whereupon the density progressively decreased to a smaller number of non-glandular trichomes per unit area on mature bracts (figure XII). The trend for clone 79 was unclear, although a slight overall decrease in trichome density during bract development appeared to exist (figure XII). The density of trichomes on clone 87 was lower than that found for clones 152 or 79, and a slight decrease in trichome density was observed as the bract matured (figure XII).
Definite trends in gland density of each gland type were evident during bract development. On each of the clones, both capitate-sessile and bulbous glands generally decreased in density as the bract developed, while capitate-stalked glands increased in density. The most prominent difference among the clones was related to the time of appearance of the capitate-stalked glands. On both clones 79 and 87, capitate-stalked glands were present in low numbers on the youngest bracts and subsequently increased in density as the bract matured. In comparison, capitate-stalked glands were not present on bracts of clone 152 until midway through bract development. These results suggest that the time of appearance of capitate-stalked glands could be characteristic for each clone, although whether this is regulated by genetic or environmental factors remains to be determined. Since these clones were grown under the same environmental conditions and the bracts from each clone were collected at the same time (1500 h) on three successive days, the effect of environmental factors, if significant, would be difficult to determine. Further studies are needed to define any environmental effects on gland presence and density at each developmental stage of bracts from different phenotypes.
In contrast to the individual gland types which showed similar patterns on each of the clones, individual cannabinoids differed from clone to clone. The characteristic cannabinoid (CBD) of both clones 79 and 87 increased in concentration during bract development, but at a different rate in each clone. Clone 152, however, differed in that the concentration of its characteristic cannabinoid ( @ 9-THC) remained stable during bract development. As stated for the glands, further studies are also needed to define to what degree environmental conditions determine the developmental trend of cannabinoid formation. Although the cannabinoid profiles differed from clone to clone, within each clone the concentrations of the individual cannabinoids followed similar patterns. An increase or decrease in the characteristic cannabinoid was generally paralleled by an increase or decrease in other cannabinoids present.
In addition to the glandular trichomes which have been shown to contain cannabinoids (Fairbairn, 1972; Turner et al., 1977, 1978), non-glandular trichomes which lack cannabinoids (Malingre et al., 1975) also are present on the plant epidermis in high numbers. As found for each of the three types of glandular trichomes, the non-glandular trichomes displayed a general trend during bract development that was similar in the three clones. Each clone maintained a relatively stable density of trichomes during early bract development with a decrease in trichome density as the bract matured. These trichomes are silicified (Dayanandan and Kaufman, 1976) and rigid, which suggests that they may serve as a defense mechanism against herbivory.
Data expressed as total numbers of capitate glands per bract indicate that new glands are initiated and develop throughout bract ontogeny. Gland density patterns for both glandular and non-glandular trichomes also demonstrated this same phenomenon. When total gland number was compared among the clones, only minor differences existed during bract ontogeny. At bract maturity, the clones demonstrated some variability; however, the general trends of total gland numbers as well as development of each gland type were similar. It would appear, therefore, that strains cannot be distinguished from each other by the number or development of glandular or non-glandular trichomes.
As found for the glands, bract cannabinoid content presented as total cannabinoids per bract suggests that cannabinoids are synthesized throughout bract development. An increase in cannabinoid content was observed regardless of the characteristic cannabinoid present in the clone. However, clone 152, a @ 9-THC-containing phenotype, had a lower total cannabinoid content than clones 79 or 87 which are characteristically high in CBD. Whether expressed as total cannabinoids per bract or as cannabinoid concentration, it was apparent that the cannabinoid profile characteristic of a strain existed in bracts at all stages of development, emphasizing the usefulness of cannabinoid profiles for taxonomic purposes.
Although the density patterns of the individual gland types showed little if any correlation with concentrations of individual cannabinoids during bract development, a comparison of the total number of capitate glands per bract with total cannabinoid content per bract does indicate that a positive correlation exists between gland number and cannabinoid content. This relationship was evident throughout bract ontogeny and was similar for each of the clones. Comparing gland number with cannabinoid content at each stage of bract development, it was possible to estimate the cannabinoid content of a single gland on the bract. The estimate (for example, 20 ng for clone 152 and 44 ng for clone 87) compared favourably with results obtained previously when the cannabinoid content of individual glands was analyzed (Turner et al., 1977, 1978). This suggests that the glands present on the bract possess the capacity to contain all of the cannabinoids found in the bract. However, this does not eliminate the possibility that small amounts of cannabinoids may also be present in other bract tissues as well.
In summary, this study has shown general trends of gland initiation and development as well as cannabinoid synthesis during bract ontogeny. Although individual gland types showed little correlation with concentrations of individual cannabinoids, a positive correlation existed for the total number of capitate glands per bract as compared with the total cannabinoid content of the bract. In addition, estimates of the cannabinoid content of individual glands suggest that the glands may contain the majority of the cannabinoids present in the bract. Further studies are in progress to determine the specific site of cannabinoid synthesis in bracts and other plant tissues.
André, C., andA. Vercruysse. Histochemical study of the stalked glandular hairs of the female Cannabisplants, using Fast Blue Salt. Planta medica, 29:361 -366, 1976.
Coffman, C., andW. Gentner. Cannabinoid profile and elemental uptake of Cannabis sativa L. as influenced by soil characteristics. Agronomy journal, 67:491-497, 1975.
Cultivation, extraction and analysis of Cannabis sativa L . By N. Doorenbos and others. Annals of the New York Academy of Sciences, 191:3- 14, 1971.
Dayanandan, P., andP. Kaufman. Trichomes of Cannabis sativa L. (Cannabaceae). American journal of botany, 63: 578 - 591, 1976.
DePasquale, A. Ultrastructure of the Cannabis sativa glands. Planta medica, 25:238 -248, 1974.
The essential oil of Cannabis sativa. By T. Malingre and others. Planta medica, 28:56-61, 1975.
Fairbairn, J. The trichomes and glands of Cannabis sativa L . Bulletin on narcotics, 24:4:29-33, 1972.
-- andJ. Liebmann. The cannabinoid content of Cannabis sativa L. grown in England. Journal of pharmacy and pharmacology, 26: 413 - 419, 1974.
Hammond, C., and P. Mahlberg. Morphology of glandular hairs of Cannabis sativa from scanning electron microscopy. American journal of botany, 60:524- 528, 1973.
Haney, A., and B. Kutscheid. Quantitative variation in the chemical constituents of marihuana from stands of naturalized Cannabis sativa L. in east-central Illinois. Economic botany, 27:193 - 203, 1973.
Latta, R., and B. Eaton. Seasonal fluctuations in cannabinoid content of Kansas marijuana. Economic botany, 29: 153--163, 1975.
Ledbetter, M., and A. Krikorian. Trichomes of Cannabis sativa as viewed with scanning electron microscope. Phytomorphology, 25: 166- 176, 1975.
Photosynthesis and cannabinoid content of temperate and tropical populations of Cannabis sativa. By F. Bazzaz and others. Biochemical systematics and ecology, 3:15- 18, 1975.
Sharma, G. Altitudinal variation in leaf epidermal patterns of Cannabis sativa. Bulletin on the Torrey Botanical Club, 102:199-200, 1975.
Small, E., and A. Cronquist. A practical and natural taxonomy for Cannabis. Taxon, 25:405-435, 1976.
Studies on Cannabis. II. Examination of the narcotic and its related components in hemps, crude drugs and plant organs by gas-liquid chromatography and thin-layer chromatography. By, M. Fujita and others. Annual Report of the Tokyo College of Pharmacy, 17: 238 - 242, 1967.
Turner, J., J. Hemphill and P. Mahlberg. Gland distribution and cannabinoid content in clones of Cannabis sativa L. American journal of botany, 64:687 - 693, 1977.
--Quantitative determination of cannabinoids in individual glandular trichomes of Cannabis sativa L. (Cannabaceae). American journal of botany, 65: 1103 - 1106, 1978.

