ABSTRACT
Introduction
Materials and methods
Results
Cannabinoid content of leaflets
Discussion
Author: J. C. TURNER, J. K. HEMPHILL, P. G. MAHLBERG
Pages: 63 to 71
Creation Date: 1981/01/01
Gland number and cannabinoid content were quantified during ontogeny of vegetative leaves from three clones of Cannabis.Initiation of capitate-sessile and bulbous glands was found to occur uniformly during leaf development. Cannabinoids were synthesized throughout leaf development as well, but at a decreasing rate. A positive correlation was found for total capitate-sessile glands per leaf as compared with total cannabinoid content of the leaf. The data also indicated that other leaf tissues in addition to the glands may contain cannabinoids.
Of the three types of glandular trichomes present on Cannabis sativa,only two (bulbous and capitate-sessile) are present on vegetative leaves, while capitate-stalked glands are found in association with the inflorescence (Hammond and Mahlberg, 1973, 1977; Turner, Hemphill and Mahlberg, 1977, 1978). Both types of capitate glands have been implicated as major reservoirs of cannabinoids (Fujita et al., 1967; Fairbairn, 1972; DePasquale, 1974; Malingre et al., 1975; André and Vercruysse, 1976; Turner et al., 1977, 1978). The vegetative leaf, therefore, represents an experimental system in which a specific capitate gland type can be studied with relation to the cannabinoids present in the leaf.
* This research was supported by research grants to P. G. Mahlberg from the Department of Agriculture of the United States of America and the National Institute on Drug Abuse.
Previous work in our laboratory has revealed that the epidermal glandular trichomes present on Cannabisappear to be a complex and dynamic system in relation to both gland ontogeny and cannabinoid content (Hammond and Mahlberg, 1973, 1977, 1978; Turner et al., 1977, 1978). In the first part of the current study (Turner, Hemphill and Mahlberg, 1981), a positive correlation was found between the total number of capitate glands and total cannabinoids in pistillate bracts. Also, the data suggested that the glands on a bract could contain the total cannabinoid content detected in the bract (Turner et al, 1981).
The purpose of this investigation is to examine the pattern of gland distribution as well as the cannabinoid profile of vegetative leaves throughout leaf ontogeny. The results should indicate to what degree the interrelationships of glands and cannabinoids found for vegetative leaves reflect those found for pistillate bracts.
Clones
Plants of three Cannabis strains were selected and cloned (Turner et al., 1977). These clones were derived from a high @9-tetrahydrocannabinol (@9-THC) strain (152), a low @9-THC, high cannabidiol (CBD) strain (79), and a high CBD strain (87). Clones were grown under ambient greenhouse conditions.
Plant parts sampled
Selected leaves from each clone were analysed for their gland number and cannabinoid content. Centre leaflets of compound leaves of increasing length were collected from vegetative plants. Lengths included 2.5 cm leaflets (very young) to 12.5 cm leaflets (mature) at 2.5 cm intervals. Leaflet samples were collected in mid-July and early September 1979. Data presented in the figures are from the September collection.
Gas-liquid chromatography ( GLC) and scanning electron microscopy
Leaves to be analysed were collected at a given time of the day (3 p.m.) and processed as described previously (Turner et al., 1977). Analyses by GLC were performed on a Hewlett-Packard 5710A chromatograph equipped with a 3380A H-P integrator. Samples prepared for scanning electron microscopy (SEM) were examined with an ETEC Autoscan.
Gland quantification
Gland number per unit area on leaves was determined by counting glands directly on the SEM screen (Turner et al., 1977). On leaflets, counts were made at the midpoint of the leaf blade, from the midrib to the margin. Multiple counts (16 fields, totalling 1 mm2)of both the adaxial and abaxial vein as well as non-vein areas were made, and the results were averaged to provide a mean for the sample. In the current experiments, the abaxial non-vein areas on young leaves (2.5-7.5 cm) were too densely covered with non-glandular trichomes to obtain an accurate gland count. Thus, for comparative reasons, all data values for the leaflet samples were averaged without including counts from the abaxial non-vein areas. The data for each leaf length sample were calculated as glands per mm2and also as total glands per leaflet. For the estimated cannabinoid content of the individual glands, the glands counted on the abaxial non-vein areas of older leaves (10-12.5 cm) were included in the calculations.
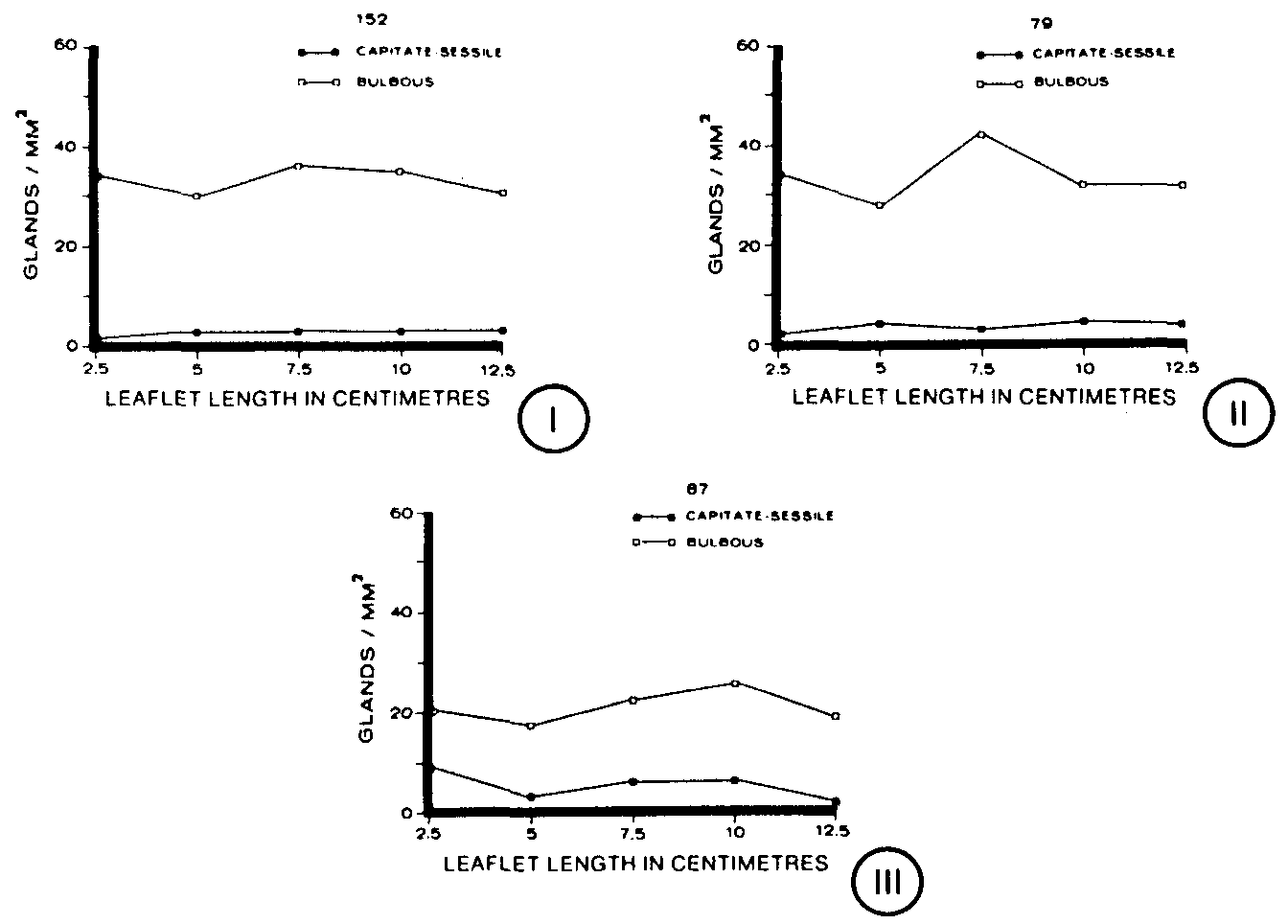
At intervals of leaf ontogeny, the number of each gland type per mm2was determined. Data collected in both July and September were essentially identical. No capitate-stalked glands were observed on any leaf samples. Capitate-sessile glands were not only present at each stage of leaf development for these clones, but also their density remained relatively constant throughout leaf ontogeny (figures I - III). Clone 152 averaged 2 capitate-sessile glands per mm2(figure I), while clones 79 and 87 averaged 5 capitate-sessile glands per mm2(figures II, III). Bulbous glands also were found to have relatively stable gland densities at all stages of leaf development for these clones (figures I-III). However, the highest gland densities of bulbous glands were found on clones 152 and 79, averaging 33 and 30 bulbous glands per mm2respectively (figures I, II). Clone 87 was found to have a lower density of bulbous glands with 21 per mm2(figure III).
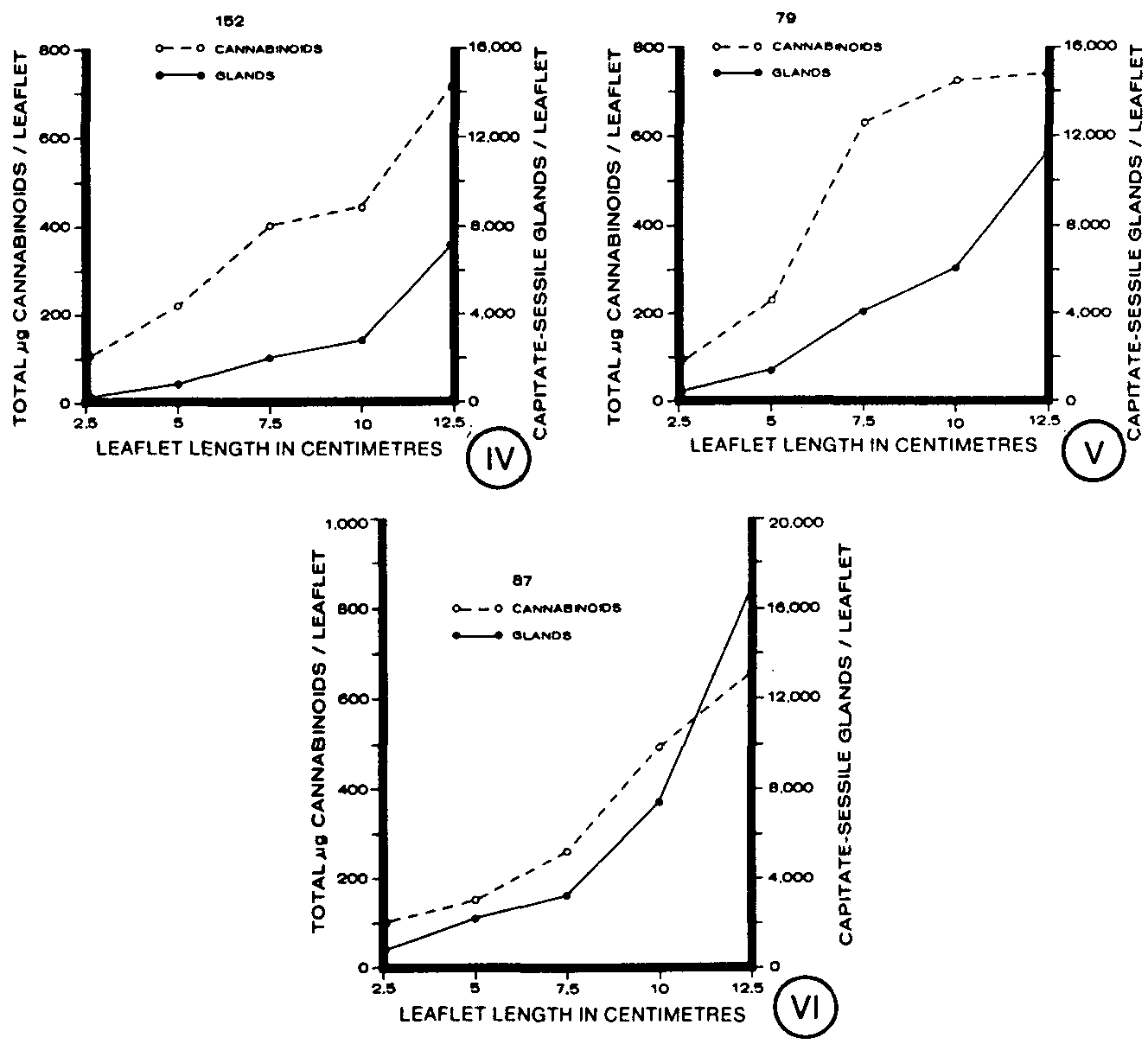
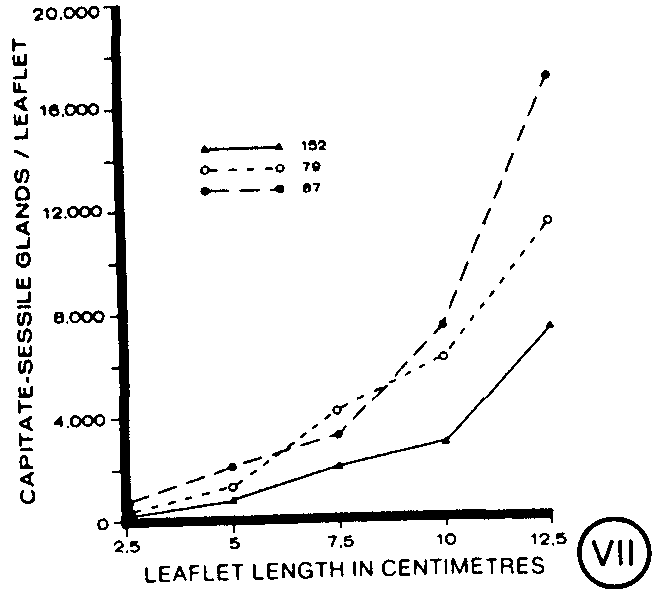
The total number of capitate-sessile glands per leaflet also was determined for these clones during leaf development (figures IV-VI). On each clone, an increase in total gland number was seen as the leaf developed, although the clones differed in the specific numbers of capitate-sessile glands found (figures IV-VI). Clone 152 had fewer glands at each development stage than clones 79 and 87 (figure VII). Clones 79 and 87 had similar numbers of glands except on mature leaflets (12.5 cm) where clone 87 had a higher number of capitate-sessile glands (figure VII).
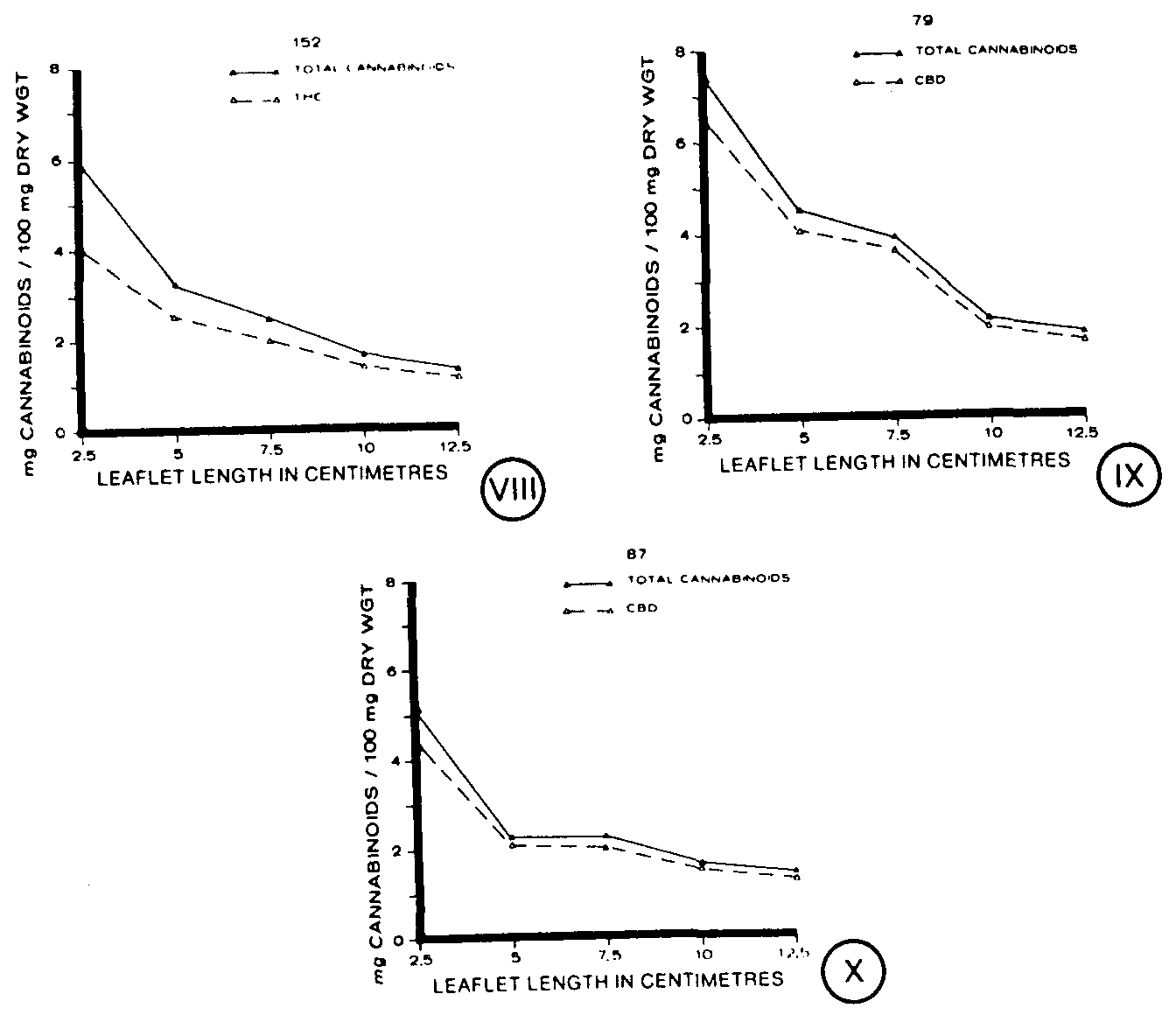
Cannabinoid concentration, on a dry weight basis, decreased in each of the clones throughout leaflet ontogeny (figures VIII-X). Although both clones 79 and 87 characteristically have high levels of CBD, clone 79 had higher levels of CBD at most stages of leaflet development than clone 87 (figures IX, X). Clone 152, a @9-THC strain, had levels of cannabinoids (figure VIII) comparable to cannabinoid levels found for clone 87 (figure X). The clones each contained concentrations of their characteristic cannabinoid at levels similar to the total cannabinoid concentration detected (figures VIII-X).
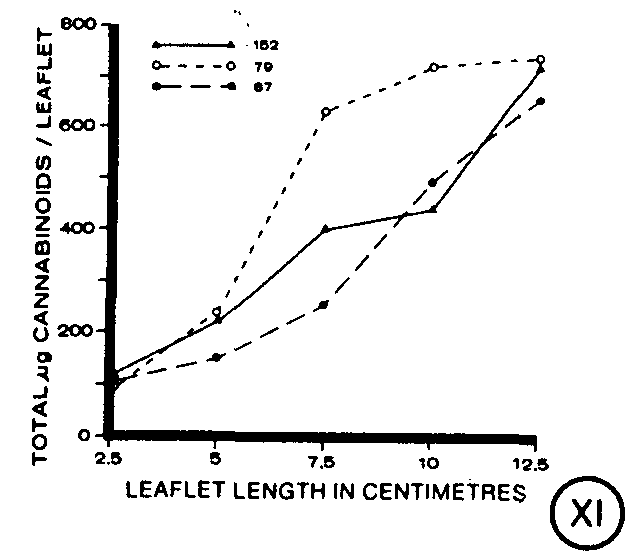
Expressed on a per leaflet basis, total cannabinoids increased during leaf ontogeny in each of the clones (figures IV-VI). The youngest as well as the most mature leaflets of the clones contained similar quantities of cannabinoids (figure XI). However, at intermediate stages, clones 152 and 87 had lower quantities of cannabinoids than clone 79 (figure XI). The increase in total cannabinoids per leaflet, in general, paralleled the increase in total capitate-sessile glands per leaflet throughout leaf ontogeny (figures IV-VI).
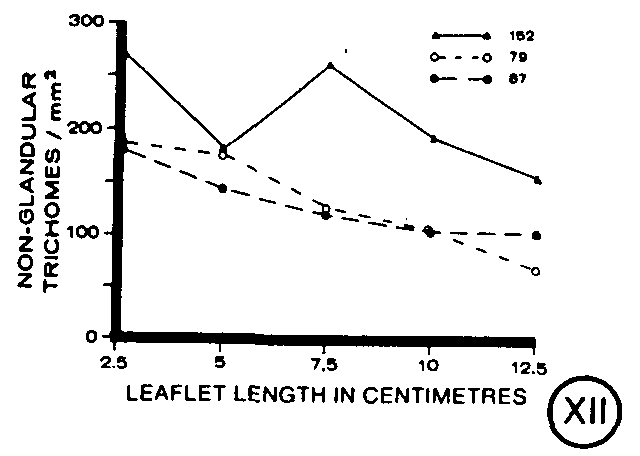
The density of non-glandular trichomes decreased slightly and at a similar rate on each clone as the leaflets enlarged (figure XII). Clones 79 and 87 were found to have comparable densities of trichomes during leaf ontogeny. However, clone 152 had approximately 25 per cent more trichomes per unit area throughout leaflet development than clone 79 or 87 (figure XII).
Vegetative leaves of Cannabisprovide an opportunity to study one capitate gland type (capitate-sessile) with relation to cannabinoid content without the presence of capitate-stalked glands. The pattern of gland distribution during leaf ontogeny differed considerably from that found during development of the pistillate bract (Turner et al., 1981). Both gland types present on the leaf, capitate-sessile and bulbous, were found to maintain a relatively constant density throughout leaf development on each of the clones. In comparison, both capitate-sessile and bulbous glands on bracts decreased in density during bract ontogeny. To what extent, if any, the appearance and increase in density of capitate-stalked glands during bract development influenced the decrease of capitate-sessile glands is unknown. However, on leaves, where no capitate-stalked glands were present, capitate-sessile glands did not decrease in density during leaf ontogeny. The considerations involved in determining patterns of gland density during leaf and bract development are unknown.
Although the density of the glands on each clone remained constant throughout leaf development, the specific numbers of each gland type varied from clone to clone. Clones 79 and 87, which are characteristically high in CBD, were found to have higher densities of capitate-sessile glands than clone 152 which contained high levels of @9-THC. Whether gland density in leaf ontogeny is influenced by the characteristic cannabinoid present in a strain is unknown. However, the presence of either @9-THC or CBD as the characteristic cannabinoid did not appear to be a major factor in determining the cannabinoid concentration found in a clone. For example, clone 152, a @9-THC-containing strain, and clone 87, a CBD-containing strain, were both found to have lower concentrations of cannabinoids than clone 79, which also is a CBD-containing strain. Although cannabinoid concentrations differed quantitatively among the clones, a trend of decreasing concentration during leaf development was found in each of the clones. In contrast, the trends for cannabinoid concentrations in developing bracts were different for each of the clones (Turner et al., 1981), Comparing cannabinoid concentrations with gland densities in these clones, it was evident that for vegetative leaves as well as pistillate bracts, there were no positive correlations between any individual gland type and any characteristic cannabinoid.
Non-glandular trichomes, reported to lack cannabinoids (Malingre et al., 1975), were found to decrease in density at a uniform rate during leaf ontogeny. This rate of decrease was slower than the rate of increase in leaf area indicating non-glandular trichomes were continually initiated during leaf development. However, fewer non-glandular trichomes were formed at each developmental stage than glandular trichomes.
A comparison of total capitate-sessile glands per leaflet with total cannabinoids per leaflet on each of the clones indicated a positive correlation as both parameters were found to increase throughout leaf ontogeny. A similar positive correlation for these parameters was also found to exist for the pistillate bracts (Turner et al., 1981). In addition, as found for the bracts, the increase in both of these parameters indicates that initiation of glandular trichomes and synthesis of cannabinoids occurs throughout leaf ontogeny.
Although these clones were similar and showed an increase in total capitate-sessile glands per leaflet and total cannabinoids per leaflet as the leaf developed, gland number and cannabinoid levels usually differed from clone to clone. Total gland number per leaflet showed the greatest variability among the clones at the most mature stage of leaf development. However, while clones 79 and 87 (both high in CBD) were relatively similar in gland number until the most mature stage, clone 152 (high in @9-THC) had fewer glands at almost every developmental stage. Whether this is due to genetic or environmental factors remains to be determined. Total amounts of cannabinoids per leaflet were generally comparable among the clones throughout leaf development, although clone 79 had higher levels at intermediate leaf stages.
As demonstrated for capitate glands on bracts (Turner et al., 1981), total cannabinoids detected per leaflet can be divided by the total number of capitate-sessile glands present per leaflet to provide an estimated amount of canna-binoids per single gland. This estimation was made for all developmental stages for each clone. When this was done for vegetative leaves, the estimated canna-binoid content per individual gland (45 ng for clone 152, 47 ng for clone 79, and 88 ng for clone 87) was higher than the cannabinoid content found for previously analysed individual glands (Turner et al., 1978). This finding differs considerably from what was found for the bract (Turner et al., 1981). On pistillate bracts, it was estimated that the gland population could contain essentially all of the cannabinoids detected in a bract. However, for vegetative leaves, it appears that the glands contain only part of the cannabinoids detected in the leaf, and, therefore, other leaf tissue may be involved in cannabinoid synthesis or accumulation.
In conclusion, this study has revealed several aspects of the interrelationship of glands and cannabinoids on Cannabis. On vegetative leaves, as on pistillate bracts, glands and cannabinoids are produced throughout organ development, although at different rates for the leaf as compared to the bract. Non-glandular trichomes are also initiated continuously during organ ontogeny. A positive correlation, as was found for the bracts, exists between total numbers of glands and total cannabinoids per leaflet. However, although glands on the bract appear able to contain almost all of the cannabinoid content of the bract, the glands on the leaf apparently contain only part of the cannabinoids detected in the leaf. It seems likely that cannabinoids are present in other tissues of the leaf. Further experiments are in progress to determine the specific site of cannabinoid synthesis in the plant.
André, C. and A. Vercruysse. Histochemical study of the stalked glandular hairs of the female Cannabis plants, using fast blue salt. Planta medica, 29:361-366, 1976.
DePasquale, A. Ultrastructure of the Cannabis sativa glands. Planta medica, 25:238-248, 1974.
The essential oil of Cannabis sativa. By Malingre, T. and others. Planta medica, 28:56-61, 1975.
Fairbairn, J. The trichomes and glands of Cannabis sativa L. Bulletin on narcotics, 24:4:29-33, 1972.
Hammond, C. and P. Mahlberg. Morphology of glandular hairs of Cannabis sativa from scanning electron microscopy. American journal of botany, 60:524-528, 1973
- - - - - Morphogenesis of capitate glandular hairs of Cannabis sativa (Cannabaceae). American journal of botany, 64: 1023- 1031, 1977.
- - - - - Ultrastructural development of capitate glandular hairs of Cannabis sativa L. (Cannabaceae). American journal of botany, 65: 140- 151. 1978.
Ledbetter, M. and A. Krikorian. Trichomcs of Cannabis sativa as viewed with scanning electron microscopy. Phytomorphology, 25: 166- 176, 1975.
Studies on Cannabis. II. Examination of the narcotic and its related components in hemps, crude drugs and plant organs by gas-liquid chromatography and thin-layer chromatography. By Fujita, M. and others. Annual report of the Tokyo College of Pharmacy, 17: 238- 242, 1967.
Turner, J., J. Hemphill and P. Mahlberg. Gland distribution and cannabinoid content in clones of Cannabis sativa L. American journal of botany, 64:687-693, 1977.
- - - - - Ouantitative determination of cannabinoids in individual glandular trichomes of Cannabis sativa L. (Cannabaceae). American journal of botany, 65: 1103- 1106, 1978.
- - - - - Interrelationships of glandular trichomes and cannabinoid content. I: Developing pistillate bracts of Cannabis sativa L. (Cannabaceae). Bulletin on narcotics, 33:2:59-69,1981.

