Introduction
Amanita species
Chemistry of the components of Amanita muscaria
The insecticidal activity of ibotenic acid and muscimol
Summary and conclusions
Acknowledgement
Author: Philip CATALFOMO , Conrad Hans EUGSTER
Pages: 33 to 41
Creation Date: 1970/01/01
Among the genera of plants, the genus Amanita (family Amanitaceae, order Agaricales, class Basidiomycetes) includes some of the most beautiful and mysterious mushrooms, showing strange chemical and pharmacological properties. Centuries have passed since observations of some of the unusual characteristics, notably, physiological effects, of these mushrooms were first documented. These characteristics include insecticidal properties, deadly toxic capabilities and the ability to cause hallucinations, narcosis and other intoxications. For over 100 years, chemists, pharmacologists and ethnobotanists, among others, have deployed great efforts in attempts to resolve the curious chemical composition of these mushrooms and to explain fully some of the peculiarities attributed to them. Despite these efforts, there remains much to be clarified.
Although several species in this taxon are scientifically interesting, of particular concern are those considered to be deadly poisonous and those known to cause hallucinations, narcosis and other centrally controlled physiological phenomena. An outline of the components known to occur in this genus follows; however, special attention will be given to the well-known fly-agaric, Amanita muscaria, because of the very curious chemistry and pharmacology of this species.
A. Poisonous Amanitas
The best known truly poisonous species- Amanita phalloides (Vaill. ex Fr.) Secr., the grünen Knollenblätterpilz-produces the bicyclic peptide toxins labelled by Wieland [ 1] as the Phallotoxin and Amatoxin-types, and which have been well described by him. The occur- rence of some of these deadly toxins has been noted also in A. verna, and by Tyler, et al. (2) in A. tenuifolia and A. bisporigera. Certain of these toxins (&alpha- and &beta-amanitin) have been reported in unrelated species, namely, Galerina autumnalis, G. marginata, and G. venenata [ 3] . As an interesting aside, Benedict et al. [ 4] were able to produce these toxins in artificial cultures of G. marginata, but attempts to duplicate this with cultures of Amanita verna were unsuccessful [ 5] . Little else is known of the over-all chemistry of the aforementioned Amanita species. It should be noted also that these toxins are not known to occur in other Amanita species [ 6,7] .
* Permanent address: School of Pharmacy, Oregon State University, Corvallis, Oregon, USA.
Note: Some ethno-botanical and anthropological aspects of the use of Amanita muscaria are found in Wasson's "Soma of the Aryans: an ancient hallucinogen?" ( Bulletin on Narcotics, volume XXII No. 3)
B. Central-Active Amanitas
Historically, Amanita muscaria (L. ex Fr.) Hooker, the red Fly-agaric, is the best known of all the Amanitas. The early history of this fascinating fungus dates back at least to the 13th century. Reviews by Tyler [ 8] , Eugster [ 9] , and recently, by Schultes [ 10] and Wasson [ 11] in this journal reveal the curiosities and mysteries surrounding this fungus.
The central-active qualities (hallucinations, narcosis and other intoxications) of Amanita muscaria have been noted for over 200 years. However, attempts to isolate and identify the active component(s), dating back almost as far, have been only partly successful. Thus the chemical knowledge to date is not sufficient to explain fully all the pharmacological actions attributed to this mushroom.
A review of the available chemico-pharmacologic information on Amanita muscaria and related species follows; highlights of the chemistry are presented here and the pharmacology of the suspected centrally acting substances will be published later.
The early chemical work dating back to Schmiedeberg and Koppe in 1869 [ 12] dealing with muscarine, the first reported substance believed to be responsible for the central-active properties of this mushroom, culminated in 1954 when Eugster, Kögl, Hardegger and co-workers fully resolved the chemistry of this relatively simple amine, which for so long defied attempts to reveal its true identity. Subsequently, Eugster and Waser submitted this now unequivocally pure compound, and its isomers, to extensive pharmacological analyses. Although muscarine could be shown to have direct action on the nervous system (it is a potent parasympathomimetic), it could not possibly be responsible for the reputed psychotropic actions of this mushroom. This conclusion was substantiated by the fact that the total muscarine content in A. muscaria is extremely low (0.0002 per cent on a fresh weight basis), which would require the ingestion of enormous quantities of the mushroom before a muscarinie effect would be elicited. Muscarine, as the responsible agent, was further precluded when it was determined that several Inocybe species, containing much larger amounts of this compound, produced toxicities unlike those of A. muscaria and never produced the hallucinations and other central effects noted for the mushroom in question [ 13,14] .
Since 1953, several reports have indicated the occurrence of suspected central-active substances in A. muscaria and related species. These were bufotenine and related indoles and the tropane alkaloids atropine, hyoscyamine and scopolamine. Lewis [ 15] was the first to report the occurrence of atropine in A. muscaria and A. pantherina. However, the yield of the alkaloid was so low that it is very doubtful that the central actions of these fungi could be attributed to its presence. Furthermore, other investigators failed to detect the presence of tropane alkaloids in the two species mentioned [ 16,17] .
Bufotenine, another central-acting substance, but only when administered parenterally, was isolated from a German specimen of Amanita citrina (A. mappa) and also reported to occur in A. muscaria and A. pantherina. Investigations of North American species revealed the presence of this indole in A. porphyria, A. tomentella, and A. citrina. It could not be detected in other Amanita species including A. muscaria and A. pantherina. Subsequent analyses of European A. citrina revealed the presence of several tryptamine derivatives in addition to bufotenine. Notable was the presence of N,N-dimethyltryptamine, a substance which is a psychotropic agent, but occurring in too low a concentration to produce any possible physiological activity of this mushroom. To date there is no indication of its existence in A. muscaria.
Interest in indolic hallucinogens was further stimulated by the discovery of psilocybin and psilocin in Psilocybe species; the latter compound being the 4-OH analog of bufotenine (5-OH-N.N-dimethyltryptamine). It has been suggested that these compounds might possibly occur in Amanita species. However, although certain indolic substances are known to occur in A. muscaria, the 4-OH tryptamine derivatives have never been repor- ted to occur therein, but they are now known to occur in Conocybe and Panaeolus species. For a review of the distribution of these substances, see [ 18] .
The most recently discovered centrally acting agents of A. muscaria are the group of isoxazole derivatives reported almost simultaneously from the laboratories of Takemoto [ 19] and Bowden [ 20] . It is interesting to note that it was the fly-killing action of these compounds, an action historically associated with A. muscaria, the fly-agaric, that attracted the attention of the Japanese group who isolated ibotenic acid from A. muscaria, A. pantherina and A. strobiliformis. Subsequently, the substance isolated by Bowden and Drysdale [ 20] , which they designated as agarin, was shown to be identical to muscimol. Prior to this time Eugster and co-workers had isolated the same compounds and the related muscazone from A. muscaria, but their findings were described only in patents and in dissertations.
The relationship between these isoxazoles and the intoxicating properties of A. muscaria was first established by Eugster and co-workers [ 21] . The extensive pharmacological testing that followed, reviewed by Waser [ 22] , established that these compounds, indeed, influence the central nervous system in animal and man. There is no doubt that muscimol is a true psychotomimetic eliciting toxic psychosis resulting in confusion, disturbed visual perception and hearing, disorientation in situation and time, etc. (Ibotenic acid and muscazone behave similarly but are somewhat less potent.) In addition, these substances potentiate the effect of certain known hypnotic/sedative agents.
The occurrence of these substances follows the peculiar pattern of limited distribution of certain compounds within the genus Amanita. A survey by Benedict, et al. [ 23] indicates that ibotenic acid and muscimol, the latter probably being an artefact generated in process, occur only in A. muscaria and A. pantherina. The chemistry and pharmacology of Amanita muscaria is discussed in the following sections. Emphasis is placed on those substances whose presence in the mushroom is unequivocal. This point is emphasized because of the numerous inconsistencies occurring in the literature dealing with the agents supposedly responsible for the intoxicating qualities of this fungus. Eventually, we hope to dispel these misconceptions and place the chemistry of Amanita muscaria in proper perspective.
Ibotenic acid: The name is derived from Ibotengu-take, the Japanese designation for A. strobiliformis (Paul.) Qu?l. Eugster and Müller [ 24] , who originally discovered this compound, provisionally called it &alpha-toxin and later [ 25] named it premuscimol. After an agreement between Eugster and Takemoto (26), the name ibotenic was retained. It has been detected in relatively few Amanita species and principally in samples from Switzerland, southern Germany, Japan and North America. (See table I.).
|
A. muscaria (L. ex Fr.) Hooker
|
0.03-0.1 % a,b
|
(25,27) |
|
A. muscaria(Fr.) S. F. Gray
|
0.0025 % a,b
|
(19) |
|
A. muscaria(Fr.) S. F. Gray var.
muscaria
|
0.18%c,d
|
(23) |
|
A. muscaria
var. formosa (Fr.) Sacc.
|
0.17%c,d
|
(23) |
|
A. muscaria
var. alba (Peck) Coker
|
0.18%c,d
|
(23) |
|
A. strobiliformis (Paul.) Qu?l.
|
0.0015%a,b
|
(28) |
|
A. pantherinn (D.C.) Fr.
|
0.021%a,b
|
(19) |
|
A. pantherina (Fr.) Secr.
|
0.46%c,d
|
(23) |
|
A. pantherina-gemmata (intermediate form)
|
0.28%c,d
|
(23) |
The nomenclature is that of the original authors although a few botanical designations are not entirely correct.
a Based on the weight of the fresh fruiting body
b Determined by actual isolation
c Based on the dry weight of the fruiting body
d Based on spot size and intensity on paper chromatograms
Other Amanita species were similarly investigated but ibotenic acid was not detected in them [ 23] ; see table 2.
|
A. gemmata (Fr.) Gill
|
A. porphyria (Fr.) Secr.
|
|
A. agglutinata (Berk. & Curt.)
|
A. rubescens (Fr.) S. F. Gray
|
|
Lloyd
|
A. silvicola Kauff.
|
|
A. aspera (Fr.) S. F. Gray
|
A. solitaria (Fr.) Secr. sensu
|
|
A. bisporigera Atk.
|
D.E. Stuntz
|
|
A. calyptroderma Atk. & Ballen
|
A. strobiliformis (Vitt.) Qu?l.
|
|
A. chlorinosma (Peck) Sacc.
|
sensu W. C. Coker
|
|
A. citrina S. F. Gray
|
A. verna
|
|
A. cothurnata Atk.
|
Vaginata fulva (Fr.) A. H.
|
|
A. flavoconia Atk.
|
Smith
|
|
A. phalloides (Fr.) Secr.
|
Vaginata livida S. F. Gray
|
All from North America except A. phalloides (Germany).
Ibotenic acid is an unstable compound; consequently, during extraction and subsequent processing large losses can occur. Also, in dried mushrooms the ibotenic acid content decreases gradually. Exact and systematic studies on the influence of varying environmental conditions on ibotenic acid content in Amanita muscaria are not available. Since Amanita muscaria is a mycorrhizal fungus, the influence of the host tree on the metabolic activity must be kept in mind also. A preliminary investigation by Eugster [ 29] makes it likely that summer mushrooms have a higher concentration than the autumn mushrooms; there were peak values up to 0.1% ibotenic acid in the fresh mushroom. It is further probable that there is an uneven distribution within the mushroom. It appears that the highest concentration is in the yellow tissue of the cap immediately below the skin. The most important active substance thus far isolated with regard to concentration is ibotenic acid. Since a medium-size fresh carpophore with a cap diameter of 10-15 cm weighs about 60-70 g, it can contain up to 70 mg of ibotenic acid. The average concentration could be about 30-35 mg when a loss through drying and possible storage is taken into consideration.
According to literature reports (41), Siberian mushroom eaters used for moderate effects 1-4 dried mushrooms and 5-10 was considered immoderate use. From this information and the current knowledge of the potency of ibotenic acid after oral ingestion (30), it follows that ibotenic acid or probably its transformation products (muscimol?) plays an important role as one of the active substances in Amanita muscaria intoxications. Unfortunately, to this time it has not been possible for us to investigate authentic specimens of the fly-agaric from Siberia. Therefore, the unequivocal proof that these too contain ibotenic acid is lacking. But, its presence is strongly suspected since samples from Europe, Japan and North America are known to contain it.
With regard to Amanita strobiliformis (whose botanic identification is not always clear) and A. pantherina, the situation is not fully understood. Benedict, et al. [ 23] could not find ibotenic acid in authenticated specimens of A. strobiliformis, in contrast to Takemoto [ 28] , but they established its presence in A. pantherina. However, Eugster et al. [ 29] could not find ibotenic acid in Swiss samples of A. pantherina. This mushroom is also listed as being hallucinogenic in the mycological literature. It is therefore desirable that a thorough and systematic investigation be conducted in order to clarify this situation, which, in reality, may merely reflect problems of nomenclature.
The procedures for the isolation and identification of ibotenic acid have been well-documented [ 23, 25] and will not be discussed here; however, certain highlights of the chemistry of this compound will be presented subsequently.
Ibotenic acid (C5H8N2O5), colourless crystals, m.p. 145° (decomposition) or 150-151° (decomposition) is an acidic, optically inactive, &alpha-amino acid with a heterocyclic substituent. It is the hydrate of ?-amino-[3-hydroxyisoxazolyl-(5)]-acetic acid (Ia) and, therefore, a glycine derivative. It is not yet known in which manner the water of hydration is bound. All attempts to prepare waterfree ibotenic acid result in a decarboxylation which under proper conditions, i.e. warming in dimethylsulfoxide, dimethylformamide, pyridine, etc., makes CO2 and water split off, and produce quantitatively muscimol (II). Waterfree esters and other derivatives of ibotenic acid can be prepared. Pure ibotenic acid is rather difficult to dissolve in cold water, but it can be recrystallized without large losses if done quickly and carefully. Impure preparations are much less stable. The easy decarboxylation makes it probable that a cooked meal of the fly-agaric cannot contain sufficient amounts of ibotenic acid to conclude that it is, per se, the true active component.
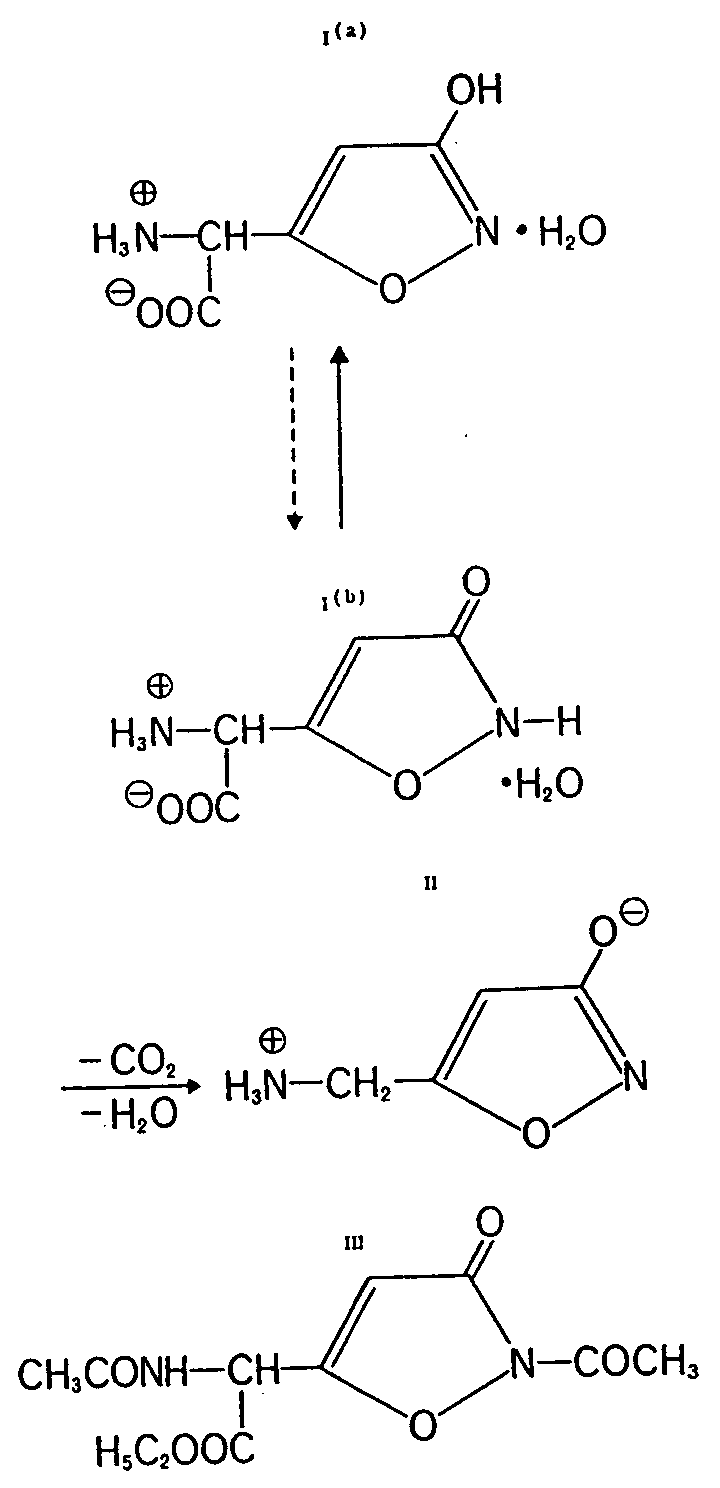
The easy decarboxylation of ibotenic acid is exceptional among natural amino acids. This simple decarboxylation is explained by the fact that trace amounts of the tautomeric lactam form (Ib) exist, which presents a vinylogous ?-keto acid as well as the strong electron withdrawing nature of the heterocyclic moiety, both of which contribute to this phenomenon. The tautomeric possibility is established in the formation of a derivative such as compound (III).
An additional characteristic of ibot enic acid is its intense yellow colour reaction with ninhydrin on paper chromatograms. The spot changes slowly to brown and then to violet after a while.
The heterocyclic group is 3-hydroxy-isoxazole. It can also be looked upon as a cyclic unsaturated hydroxamic acid. Such compounds, up to now, were unknown in nature. Of chemical significance is the easy hydrogenolysis of the N-O bond in the heterocycle, thereby producing a very reactive derivative of an &alpha-amino ketone which, depending upon conditions, can be transformed into different derivatives (i.e. dimerization to pyrazine derivatives) structurally similar to a &upsih-aminobutyric acid derivative.
Details of the wide variety of chemical reactions of ibotenic acid have been well documented [ 9] .
The interesting physiological activity of ibotenic acid stimulated several chemical syntheses of this compound [ 9] .
Muscimol:(C4H6O2N) colourless crystals, m.p. 155-156° (hydrate), 174-175° (waterfree) is the enol betaine of 5-aminomethyl-3-hydroxyisoxazole (II). Its salt nature renders it very soluble in water but only sparingly soluble in organic solvents, i.e. alcohol. Its colour reaction with ninhydrin in paper chromatograms is intense yellow and exhibits the same colour changes as ibotenic acid.
Muscimol can be found in all Amanita species in which ibotenic acid occurs. However, since it is easily derived from ibotenic acid through the loss of water and CO2, which can occur during extraction or on paper chromatograms, one cannot say positively that it is a genuine compound in the mushroom. In biological tests, muscimol is at least 5 times more active than ibotenic acid [ 30] . There results the interesting case where a simple chemical reaction (decarboxylation), which can occur during storage, in process, or in the body itself, renders a compound that is considerably more potent than the original form.
The chemical behaviour of muscimol is similar to ibotenic acid since it is also a derivative of 3-hydroxyisoxazole. Several synthetic chemical methods have been worked out to produce muscimol, as well as numerous analogs and homologs. This compound is therefore readily available if needed [ 9] .
Muscazone: This heterocyclic substituted glycine derivative is to date found only in the European fly-agaric (25). It is a neutral amino acid with the empirical formula C5H6O4N2 and produces colourless crystals from water. Also, this compound is optically inactive and is, therefore, as ibotenic acid, racemic in character. Its structure (IV) indicates it to be a 2(3H)-oxazolone derivative (&alpha-amino-[2(3H)-oxazolonyl-(5)]-acetic acid). Such amino acids have been unknown in nature to date.
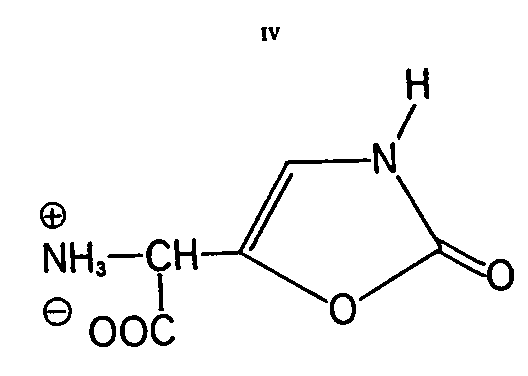
As with compounds I and II, its ninhydrin reaction is intense yellow but the colour change to violet is somewhat slower.
In chemical reactions muscazone behaves differently from ibotenic acid, with which it is a structural isomer if water of crystallization is disregarded [ 31] . The isomer is derived formally through interchange of the lactam function in Ib.
Chemically, this conversion has not yet been successful; but, it can be achieved through radiation with high energy ultra-violet light [ 32] . It is possible that in the organism muscazone is formed from ibotenic acid. Supporting this idea is the racemic character which would be hard to understand otherwise. But, it cannot be completely excluded that muscazone is an artefact generated during extraction and processing of ibotenic acid.
In pharmacological investigations, muscazone was found to be less active than either ibotenic acid or muscimol.
An insecticidal activity, especially a fly-killing one, has been traditionally associated with Amanita muscaria - the fly-agaric (Fliegenpilz). Several authors have indicated this activity in the common names of this mushroom (English: fly-agaric or bug agaric; French: tue-mouche; Russian: muchomor; Italian: moscario). However, according to Wasson, derivation of term Fliegenpilz may be explained by other reasoning. In reality, the over-all insecticidal activity of Amanita muscaria is relatively weak. Yet, it was just this insecticidal activity which stimulated Takemoto, et al. in Japan [ 19] and Bowden in England [ 20] to isolate ibotenic acid and muscimol, resp. However, Eugster, et al. [ 21] were attracted to these compounds because of their narcotic potentiating nature.
Recent quantitative investigations on the pure substances* showed that in "contact" experiments with flies no effect could be observed and, when "fed", only minimal activity was noted. Mosquitoes were somewhat more susceptible. The results are summarized in tables 3 and 4.
Muscarine: (for extensive review see Eugster [ 9] ) Muscarine occurs in the fresh fly-agaric, according to more recent data, up to 0.0002-0.0003% calculated as its chloride. The literature published before 1954, in which usually much higher concentrations were reported, should be disregarded since most likely the results are in error because they were derived from assays using impure muscarine as the reference standard.
Conducted in the Abteilung Angewandte Chemie, J. R. Geigy AG, Basel, 1965. We are indebted to Dr. H. Gysin and Dr. R. Gasser for these results.
|
Percent dead flies |
||
|---|---|---|
| 20 |
mg/filter disc
|
|
|
muscimol
|
11 | |
|
ibotenic acid
|
25 | |
|
Dimetila
|
94 | |
| 40 |
mg/filter disc
|
|
|
muscimol
|
45 | |
|
ibotenic acid
|
28 | |
|
Dimetila
|
100 |
a Compound administered with honey and water.
|
Per cent inactive after 5 hours |
||
|---|---|---|
| 10 |
mg/test plate
|
|
|
muscimol
|
90-100
|
|
|
ibotenic acid
|
50-100
|
|
| 20 |
mg/test plate
|
|
|
muscimol
|
100 | |
|
ibotenic acid
|
90-100
|
|
| 1 |
mg/test plate
|
|
|
Diazino
|
100 |
Furthermore, no attention was given to the presence of acetylcholine in the mushroom. The latter compound no doubt caused the results of biological tests to be inaccurate. In addition, the older reports on muscarine in Amanita pantherina, Boletus luridus, Russula emetica, etc., must be re-evaluated since they are based on the same aforementioned inconsistencies. On the other hand, muscarine is not only quite common in the genus Inocybe(and also in certain Clitocybe species) but it also occurs in astonishingly high concentrations. The occurrence of muscarine based on newer analyses is summarized in table 5.
Paper chromatographic and biological determinations were recently carried out by Tyler, et al. [ 33,34,35] ; refer also to the summary in [ 9] .
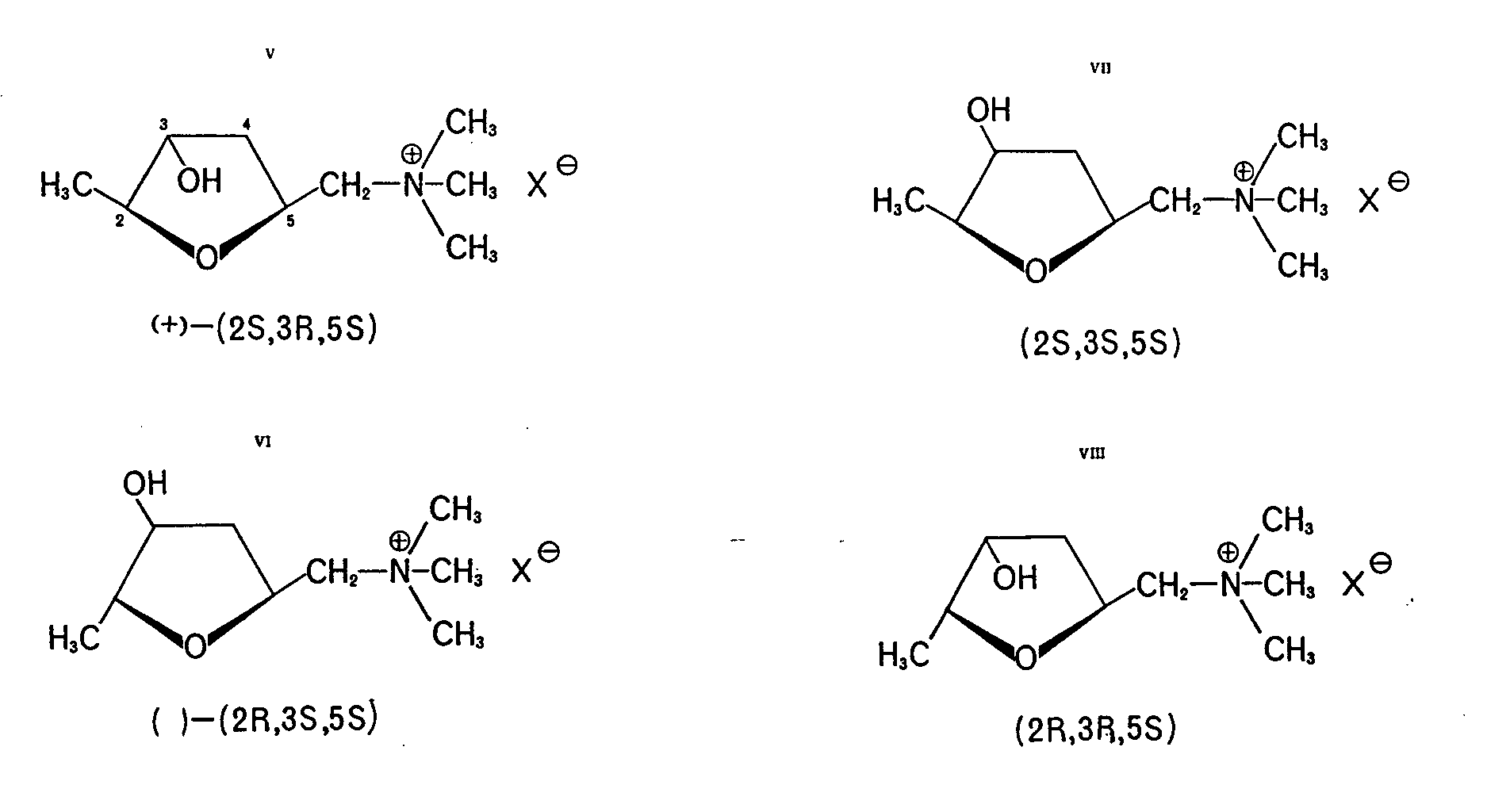
Muscarine has structure V; this means that in the mushroom the species present has the (2S,3R,5S) configuration. The enantiomeric (mirror image) molecule, which was synthesized, proved to be less potent in biological tests as were also other isomers, namely, allo-muscarine (VI), epi-muscarine (VII) and epiallo-muscarine (VIII).
The muscarines are quaternary ammonium salts, derivatives of ?-hydroxy-tetrahydrofuran, and highly soluble in water (except for salts of certain complex anions). They are stable and not destroyed by cooking
|
Amanita muscaria (L. ex Fr.) Hooker
|
0.0002-0.0003 % a b
|
I.
Stuntz 2147
|
0.476 % c d,
|
|
Inocybe Patouillardi Bres.
|
0.037 % a b
|
0.07-0.18 % c e
|
|
|
I.
fastigiata (Schff. ex Fr.) Qu?l
|
0.01 % a b
|
I.
Stuntz 3365
|
nilc e
|
|
I. umbrinaBres.
|
0.003 % a b
|
I.
griseolilacina Lange
|
0. 835 % c d,
|
|
0.27 % c e
|
0.17 % c e
|
||
|
I.
Bongardi (Weinm.) Qu?l.
|
nil.a b
|
I.
Stuntz3399
|
0.116 % c d,
|
|
I.
lilacina(Boudier) Kauffman
|
0.25-0.31 % c d,
|
0.05 % c e
|
|
|
0.38 % c e
|
I.
Stuntz1774
|
0.105 % c d,
|
|
|
I.
obscuroidesOrton (Stuntz 3761)
|
0.32-0.52 % c d,
|
0.06 % c e
|
|
|
0.11-0.80 % c e
|
I.
Stuntz 2907
|
0.28-0.30 % c e
|
|
|
I.
sororia Kauffman
|
0.26-0.35 % c d,
|
I.
Stuntz 3983
|
0.12-0.13 % c e
|
|
0.13-0.28 % c e
|
I.
lacera (Fries) Qu?let
|
0. 846-1.00 % c d,
|
|
|
I.
nigrescens Atkinson
|
nil c d, nil c e
|
0.08 % c e
|
|
|
I.
napipesLange
|
0.23-3.15 % c d,
|
I.
griseoscabrosa (Peck) Earle
|
nilc e
|
|
0.71 % c e
|
I.
pallidipes Ellis & Everhart sensu Kauff.
|
0.16c e
|
|
|
I.
picrosmaStuntz
|
0.005 % c d, nil c e
|
I.
cinnamomea A. H. Smith
|
0.251 % c d,
|
|
I.
Kauffmanii A. H. Smith
|
0.486 % c d
|
0.03 % c c
|
|
|
I.
Stuntz4292
|
0.144 % c d
|
I.
mixtilis(Britz.) Saccardo
|
1.33 % c d,
|
|
I.
Stuntz 1790
|
0.255 % c d, nil c e
|
0.10 % c e
|
|
|
I.
Stuntz 1838
|
0. 158 % c d,
|
I. xanthomelas Boursier & Kühner
|
0.09 % c d, nil c e
|
|
0.03 % c e
|
I. praetervisaQu?let
|
0.107 % c d
|
|
|
I.
terriferaKühner
|
0.269 % c d,
|
I.
albodiscaPeck
|
0.003 % c d, nil c e
|
|
0.01 % c e
|
I.
oblectabilis(Britz.) Saccardo
|
0.317 % c d
|
|
|
I.
geophylla (Fries) var. geophylla P.
|
fma. decemgibbosa Kühner
|
||
|
Karsten
|
0.259 % c d,
|
I.
Stuntz3832
|
0.161 % c d
|
|
0. 16 % c e
|
I.
decipientoides Peck
|
0.782 % c d
|
|
|
I.
pudica Kühner
|
0.117 % c d,
|
I.
Stuntz 1540
|
1.98 % c d,
|
|
0.12-0.17 % c e
|
|||
|
I.
agglutinata Peck
|
0.31-0.32 % c e
|
I.
hirsuta var.
maxima A. H. Smith
|
nilc e
|
|
I.
Stuntz 3691
|
0.08 % c e
|
I.
Stuntz1187
|
0.10 % c e
|
|
I.
olympiana A. H. Smith
|
0.336 % c d
|
I.
Stuntz 4291
|
0.11 % c e
|
|
I.
subdestricta Kauffman
|
0.421 % c d
|
I.
subexilis (Peck) Saccardo
|
nilc e
|
|
0.22 % c e
|
Clitocybe dealbata (Sow. ex Fr.) Kummer
|
0.15 ± 0,04 % c b e
|
|
|
I.
gausapataKühner
|
0.438 % c d
|
C. rivulosa(Pers. ex Fr.) Kummer
|
0.013 % f b
|
a Based on fresh weight of fruiting body.
b Determined by actual isolation.
c Based on the dry weight of fruiting body.
d Determined by biological tests.
e Base on spot size and intensity on paper chromatograms.
f Based on dry weight of mycelium.
contrary to some opinions. However, through pouring off of the water in which they were cooked, large amounts of muscarine are most likely lost.
Muscarine and ibotenic acid are not evenly distributed throughout the fruiting body of the fly-agaric. The red skin and probably also the yellow tissue immediately below contain more of these compounds than other parts. Therefore, the earlier popular recommendation of "peeling" before eating is now justified [ 40] .
The isomers of muscarine are not easy to distinguish from one another because of their similar chemical characteristics. Recent investigations showed that natural muscarine can be accompanied by smaller amounts of isomers VI, VII, and VIII* in the fly-agaric as well as in Inocybespecies [ 36, 37] . It appears possible that certain Inocybe species produce more muscarine isomers than muscarine itself. If in mushrooms the enantiomeric (-)-(2R,3S,5R)-muscarine also occurs is a question which cannot be answered yet because of experimental difficulties. These results do show, however, that if only biological testing is used to determine the muscarine content then erroneous conclusions may result since the possible presence of other isomers would be overlooked.
The exceptional specificity of muscarine activity stimulated the production of several chemical syntheses of muscarine, its isomers, analogs and homologs. The small and compact molecule possesses an easily visible stereochemistry. Therefore it is readily understandable that it is a very good system to study structure-activity relationships. In this connection refer to review (9).
In summary, muscarine is only a minor component in Amanita muscaria. The central activity of fly-agaric presumably does not involve muscarine.
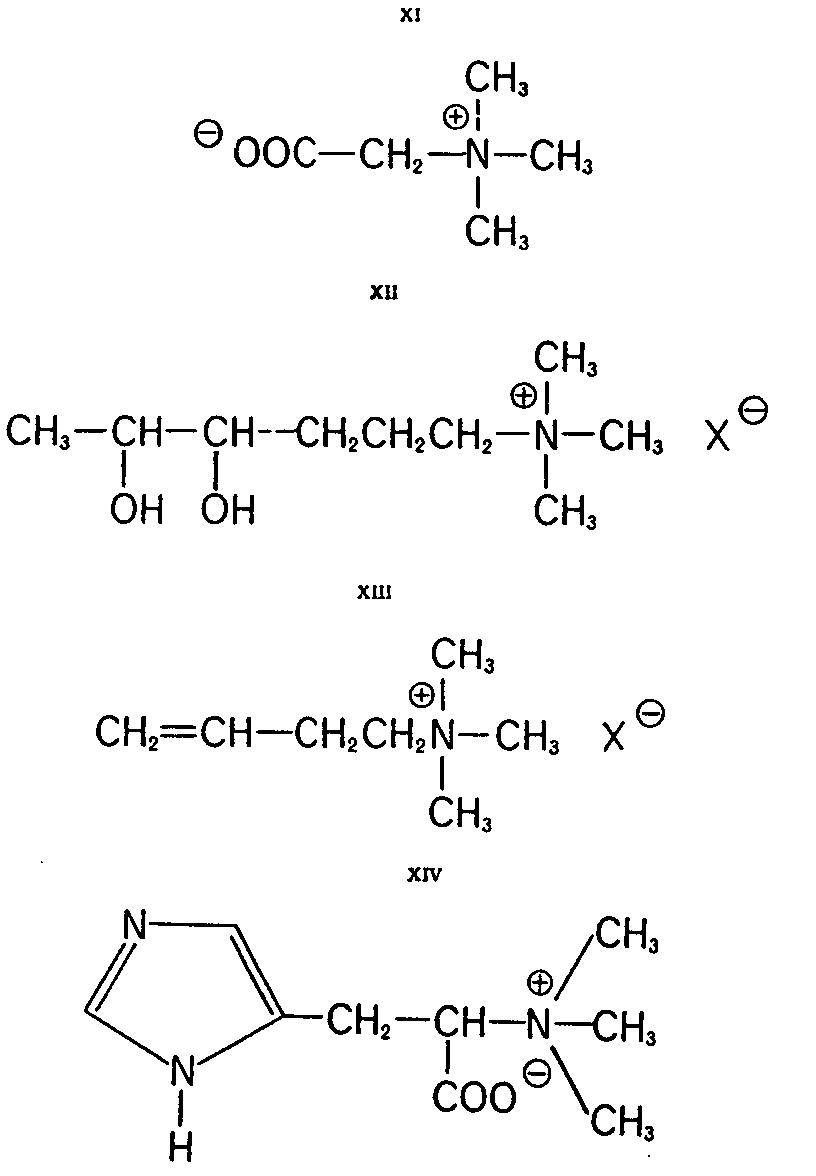
Other Compounds of Amanita muscaria (9) Additional quaternary compounds that have been reported to date from the fly-agaric include: choline (IX), acetylcholine (X), betaine (XI), "muscaridine" (XII), butenyl-trimethyl ammonium salt XIII and hercynine (XIV).
Choline (IX) appears to occur in large quantities but quantitative data are not available. The compounds X, XI, XII and XIII were isolated in very small amounts

Absolute configurations are known to date only for IV and VI. by Kögl, Salemink, et al. (9). The name "muscaridine" is an unfortunate choice for compound XII. It should not be confused with Kobert's conception for the central active principle of the fly-agaric, namely, pilzatropine-a descriptive term indicating an activity and not a specific known entity!
To date, it has not been proved that any of these compounds contribute significantly to the mushroom toxicity.
Other substances isolated from the fly-agaric include: (-)-R-4-hydroxypyrrolidone-(2) (XV), uracil (XVI), hypoxanthine (XVII), xanthine (XVIII), adenosine (XIX), the carboline derivative XX and &beta-D-n-butylglyco-pyranoside (XXI).
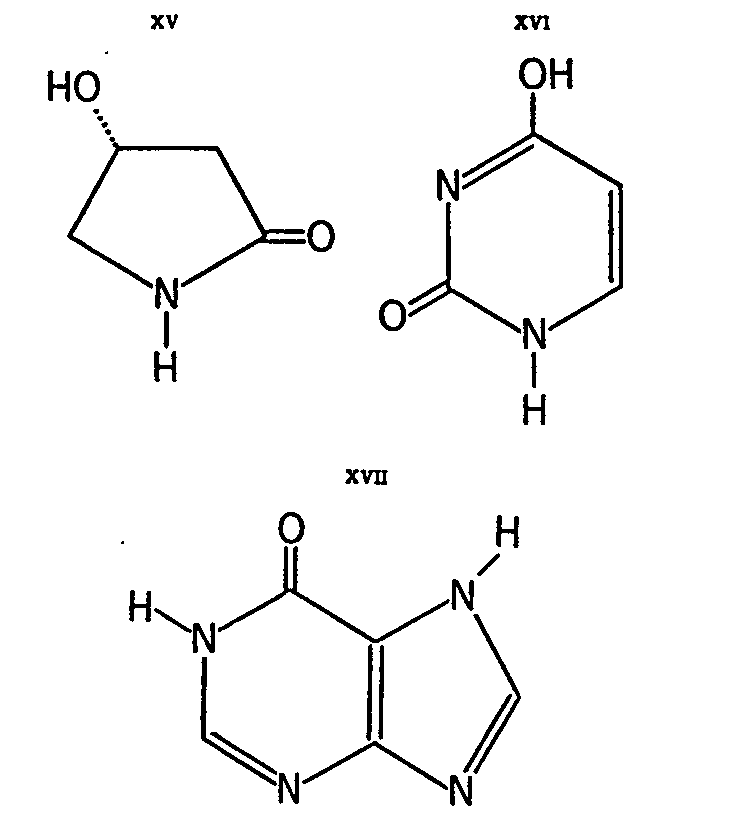
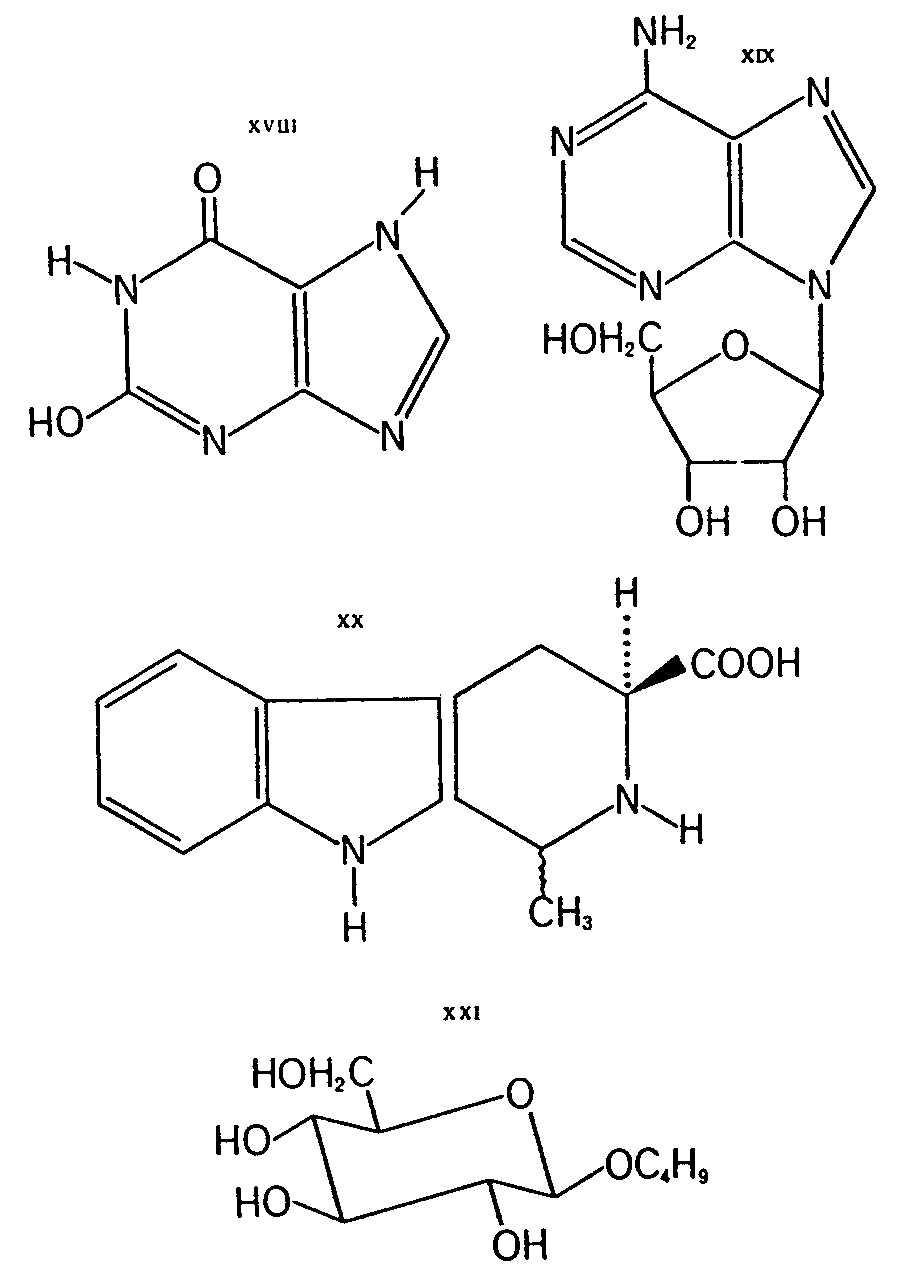
Compound XV exists only in trace amounts in the fly-agaric (quantitative data are not available), but it merits consideration as a derivative of &upsih-aminobutyric acid (GABA) and as such in connection with ibotenic acid and muscimol. Little is known about its biological activity. It was isolated from fractions which showed activity that was antagonistic to certain known narcotics [ 38] . The remainder of the substances are probably not genuine secondary metabolites and most likely they are not involved in the pharmacological activities of the mushroom.
Pigments. In the chemical literature the red pigment of the cap of the fly-agaric is called muscarufin by Kögl and Erxleben [ 39] and structural elucidation stated it to be a terphenylquinone derivative (XXII).
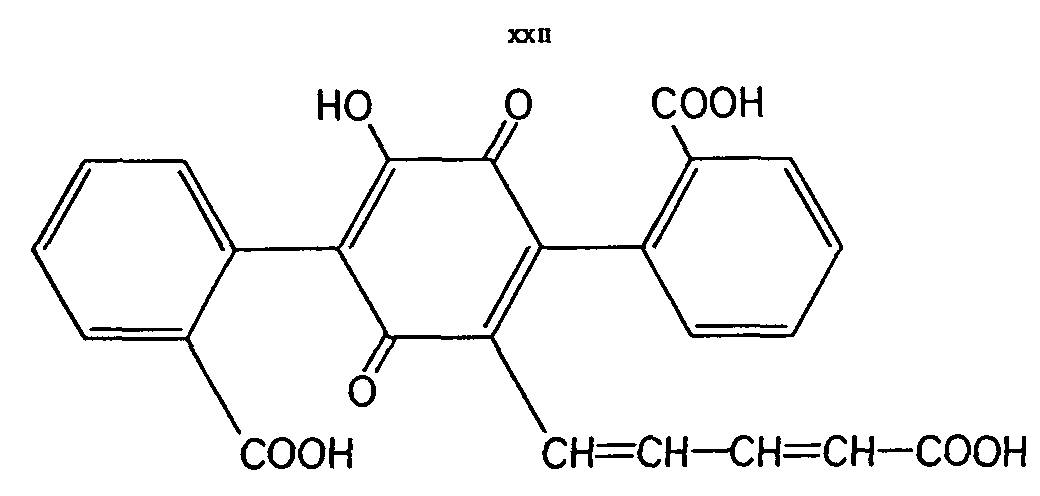
There are several indications that this is not correct. A pigment of this type could not be verified, nor did chemical reactions lead to compounds with the charac teristics of muscarufin. In reality, the yellow and red pigments of Amanita muscaria are a complicated mixture of very labile compounds which without doubt cannot represent terphenylquinones. The characteristics rather show that they belong to a different class of compounds [ 9] .
A complete list of compounds isolated to date from Amanita muscaria, but not discussed here, which includes organic acids, esters, sugars, sugar alcohols, amines, amino acids, nucleotides, etc. are given in [ 9] .
Our present knowledge of the chemistry of Amanita muscaria cannot explain fully the physiological actions stimulated by the ingestion of adequate amounts of this fungus. To accommodate the wide range of central actions has not been an easy task, and it is unlikely that only one compound will prove to be the causative agent. No doubt, ibotenic acid and/or muscimol play a prominent role, but the pharmacological data to date will not allow us to conclude that these compounds are the sole active agents. It remains to be determined whether or not the total "toxicity" caused by this mushroom is due to as yet undiscovered substances or if the combined actions of several substances ultimately prove to be the answer.
On the other hand, several reports in the older literature dealing with the chemistry and pharmacology of this mushroom present what now must be considered erroneous conclusions with regard to the role played by certain substances in the intoxicating properties of Amanita muscaria. Indeed, in certain instances, it remains highly questionable if the substance reported to be responsible for the physiological activity actually occurs in the mushroom. Specifically, the following points come to light.
Muscarine does occur in A. muscaria but in such a low concentration that it is untenable to consider that it is the principal agent responsible for the physiological action attributed to this mushroom. Moreover, the results of careful pharmacological testing preclude this compound as the centrally active agent.
Pilzatropine, a descriptive term coined by Kobert to express an action, is sometimes believed to be a specific chemical entity. It is not a chemical compound but merely a term describing the similarities between atropine poisoning and that caused by the fly-agaric.
Tropane alkaloids, in very low concentrations, have been reported to occur in this mushroom but not to the complete satisfaction of interested scientists since several studies have failed to reveal these compounds in specimens from Europe and North America. Although it is possible in principle that tropane alkaloids may occur in A. muscaria, definitive proof must be based on further thorough investigations.
A similar situation involves the occurrence of bufotenine reported to occur in Amanita muscaria. Several recent studies have failed to reveal this compound in the fly-agaric. In some toxicology texts it is claimed that bufotenine is the hallucinogenic agent in A. muscaria. This is an error not only because the compound does not exist in this mushroom but also because of the well established fact that bufotenine is inactive when administered orally.
We hope that we have placed these inconsistencies in proper perspective and further hope that they cease to be perpetuated in future literature. In the meantime, work continues in order that eventually we may establish which chemical entities are responsible for the physiological activities of this fascinating but perplexing red mushroom- Amanita muscaria.
We are grateful to the American Foundation for Pharmaceutical Education for a Gustavus A. Pfeiffer Memorial Research Fellowship held by one of us (P.C.) during 1969-1970.
T. Wieland, Science, 159, 946 (1968)
002V. E. Tyler Jr., R. G. Benedict, L. R. Brady and J. E. Robbers, J. Pharm. Sci., 55, 590 (1966)
003V. E. Tyler Jr., L. R. Brady and R. G. Benedict, Lloydia, 26, 154 (1963)
004R. G. Benedict, V. E. Tyler, Jr., L. R. Brady and L. J. Weber, J. Bacteriol., 91, 1380 (1966)
005P. Catalfomo, unpublished data, 1968
006P. Catalfomo and V. E. Tyler, Jr., J. Pharm. Sci., 50, 689 (1961)
007S.S. Block, R. L. Stephans and W. A. Murrill, J. Agr. Food Chem., 3, 584 (1955)
008V. E. Tyler, Jr., Am. J. Pharm., 130, 264 (1958)
009C. H. Eugster, Fort. Chem. Organ. Naturst., 27, 261 (1969)
010R. E. Schultes, Bull. Narcotics, 21, No. 3, 3 (1969)
011R. G. Wasson, ibid., 22, No. 3 (1970)
012O. Schmiedeberg and R. Koppe, "Das Muscarin, das giftige Alkaloid des Fliegenpilzes" Leipzig: F.C.W. Vogel 1869
013C. Fahrig, Arch. Exp. Path. Pharm., 88, 227 (1920)
014W. Mecke, ibid., 175, 23 (1934)
015B. Lewis, South African Med. J., 29, 262 (1955)
016L. R. Brady and V. E. Tyler, Jr., J. Am. Pharm. Assoc., Sci. Ed. 48, 417 (1959)
017C. A. Salemink, J. W. ten Broeke, P. L. Schuller and E. Veen, Planta med., 11, 139 (1963)
018R. G. Benedict and L. R. Brady, "Fermentation Advances", Academic Press, N.Y., 63 (1969)
019T. Takemoto, T. Nakajima and R. Sakuma, Yakugaku Zasshi, 84, 1233 (1964)
020K. Bowden and A. C. Drysdale, Tetrahedron Letters, 12, 727 (1965)
021G. F. R. Müller and C. H. Eugster, Helv. Chim. Acta, 48, 910 (1965)
022P. G. Waser , Ethnopharmacologic Search for Psychoactive Drugs, D. H. Efron, Bo Holmstedt and N. S. Kline, ed., U.S. Dept. Health, Educ. Welf., Publ. Health Serv. Publ. No. 1645, Washington D.C. 1967
023R. G. Benedict, V. E. Tyler, Jr. and L. R. Brady, Lloydia, 29, 333 (1966)
024G. F. R. Müller, Dissert. Univ., Zürich, 1961
025R. Good, G. F. R. Müller and C. H. Eugster, Helv. Chim. Acta, 48, 927 (1965)
026C. H. Eugster and T. Takemoto, Helv. Chim. Acta, 50, 126 (1967)
027C. H. Eugster, G. F. R. Müller and R. Good, Tetrahedron Letters, 1965, 1813
028T. Takemoto, T. Yokobe and T. Nakajima, Yakugaku Zasshi, 84, 1186 (1964)
029C. H. Eugster, unpublished data, 1964
030W. Theobald, O. Buch, A. Kunz, P. Krupp, E.G. Stenger and H. Heimann, Arzneimittelforsch., 18, 311 (1968)
031R. Reiner and C. H. Eugster, Helv. Chim. Acta, 50, 128 (1967)
032H. Göth, A. R. Gagneux, C. H. Eugster and H. Schmid, Helv. Chim. Acta, 50, 137 (1967)
033J. K. Brown, M. H. Malone, D. E. Stuntz and V. E. Tyler, Jr., J. Pharmaceut. Sci., 51, 853 (1962)
034M. H. Malone, R. C. Robichaud, V. E. Tyler, Jr. and L. R. Brady, Lloydia, 24, 204 (1961)
035M. H. Malone, R. C. Robichaud, V. E. Tyler, Jr. and L. R. Brady, Lloydia, 25, 231 (1962)
036C. H. Eugster and E. Schleusener, Helv. Chim. Acta, 52, 708 (1969)
037P. Catalfomo and C. H. Eugster, Helv. Chim. Acta, 53, 848 (1970)
038T. Matsumoto, W. Trueb, R. Gwinner and C. H. Eugster, Helv. Chim. Acta, 52, 716 (1969)
039F. Kögl and H. Erxleben, Liebigs Ann. Chem., 479, 11 (1930)
040C. H. Eugster and G. F. R. Müller, Helv. Chim. Acta, 42, 1189 (1959); K. Bowden, A. C. Drysdale and G. A. Mogey, Nature, 206, 1360 (1965)
041R. G. Wasson, Soma. Divine Mushroom of Immortality, Harcourt, Brace and World, Inc. N.Y. (1968), p. 233 ff.


