Experimental technique
Results
Conclusions
Summary
Author: A. de PASQUALE, G. TUMINO, R COSTA de PASQUALE
Pages: 27 to 40
Creation Date: 1974/01/01
The electron scanning microscope is of great use in pharmacognosy because it enables valuable contributions to be made to the study of the micromorphology of the superficial tissues of drugs. The considerable resolving power of this instrument and its great depth of field give it clear advantages over the light microscope.
Morphological micro-characteristics may be of great importance both for drug identification, particularly in cases in which the other characteristics are insufficient, and for plant taxomony.
The observation of epidermes by means of a scanning microscope, which reveals the finest details of the cuticle, stomata, trichomes, etc., is of particular value in the case of Cannabis sativa L., because it permits very precise comparisons of the characteristics of such elements, some of which may assume a quality of specificity. This plant, in fact, tends to deviate from the original strain according to its environment; moreover, plants obtained from seeds from different sources and grown in the same region tend to assume similar characteristics, not only from the morphological standpoint, but also as regards resin content and composition.
The micromorphology of hemp may also be of legal importance, because the observation of certain special morphological features, such as glandular, cystolith and covering hairs, can assume probative value, in conjunction with chemical and chemico-physical analyses and biological tests.
The trichomatous formations of hemp have generally been described as belonging to the following three types: (1) capitate glandular hairs; (2) unicellular cystolith hairs; and (3) unicellular covering hairs of varying form and size in the different parts of the plant.
In a previous study [ [ 1] ] , using a light microscope, we observed how, in addition to the capitate glandular hairs, there were other hairs - sessile hairs or hairs with very short stalks - present on the bract covering the female flower - referred to as the "perigonal" bract [ [ 1] ] , on the tepals and anthers of male flowers, and on the lower surface of leaves.
These observations were confirmed during our successive research using the transmission electron microscope and the scanning electron microscope [2,3]. Only Fairbairn [ [ 5] ] has recently detected, on the lower surface of the leaves and bracts, and on the anthers, large sessile glands.
In the course of other earlier research [ [ 4] ], we had demonstrated how the active cannabinoid elements are present in all the aerial parts of the female hemp plant, and also in small quantities in the roots. We nevertheless felt that it would be useful to examine under the scanning electron microscope the epidermis of the stalk, the leaf limba, the petiole and the floral and perigonal bracts at the various stages of development of inflorescence and infructescence, in order to identify any relationships that might exist between superficial glandular formations and resin production.
The various fresh parts of the female plants of Cannabis sativa L., derived from seeds from Amindeon in Greece and grown at Messina, were fixed immediately after removal, using 4 per cent glutaraldehyde buffered at pH 7.2 by Sorensen's phosphate buffer solution (0.05 M), then washed for a long time in water, and finally dried for lyophilization. The purpose of lyophilization is to prevent, as far as possible, distortions due to dessication and vacuum conditions, particularly in the case of the most frail samples with a high water content. The specimens were subsequently coated with gold in a JEE-4B Vacuum Evaporator-Jeol. Observations were made with a Jeol-JSM-S1 scanning electron microscope.
Two Zones are apparent in the stalk: ( a) the lower part; and ( b) the green tips of the stalk and branches.
In the lower part and in the intermediate part, whose structure is secondary, the thin layer of cork which replaces the epidermic tissue appears to be formed of cells which are mostly rectangular (fig. 1), regular and covered with suberous lamellae. Only in places, where suberification was not complete, was it possible to detect very occasional unicellular, long, slender covering hairs with a verrucose surface; no glandular hair was detected.
In the younger parts of the stalk and branches, whose structure is primary, the epidermic tissue is composed of rectangular or isodiametric cells, with regular, straight contours and occasional stomata. The youngest cells (fig. 2) are covered with a thin cuticle; on the older cells (fig. 3), it is possible to detect irregular layers of cutin, which appears to accumulate mainly on the guard cells of the stomata, on the related cells, and on the cells surrounding the base of the cystolith hairs. Along the lines of contact between two neighbouring cells, the cutin is often arranged in a string-of-beads pattern.
The very numerous covering hairs are unicellular, with verrucose surfaces, vermiform, of varying sizes, straight or curved, and in some cases bent in the manner of a knee: some hairs have branches which anastomose with neighbouring hairs. Cystoliths (fig. 4) are infrequent, short, isolated or grouped in clusters of three or four. We observed very infrequent sessile glands approximately 45-55 µm in diameter.
The petiole has, running parallel to the axis, longitudinal protrusions along which the epidermis appears to be composed of rectangular, slender cells elongated in the direction of the axis itself, and it has conical unicellular hairs, with verrucose surfaces, some of which are very thick and long, and others distinctly smaller. In the hollows (fig. 5), between the above-mentioned protrusions, polygonal cells, covered with a striated cuticle, can be observed, with infrequent stomata and very numerous short hairs (fig. 5), which are unicellular, conical, straight or curved, with smooth or verrucose surfaces. In the hollows it is also possible to detect more numerous sessile glandular hairs than in the stalk, swollen, 40-60 µm in diameter, and others which are less developed, considerably smaller and 15-20 µm in diameter.



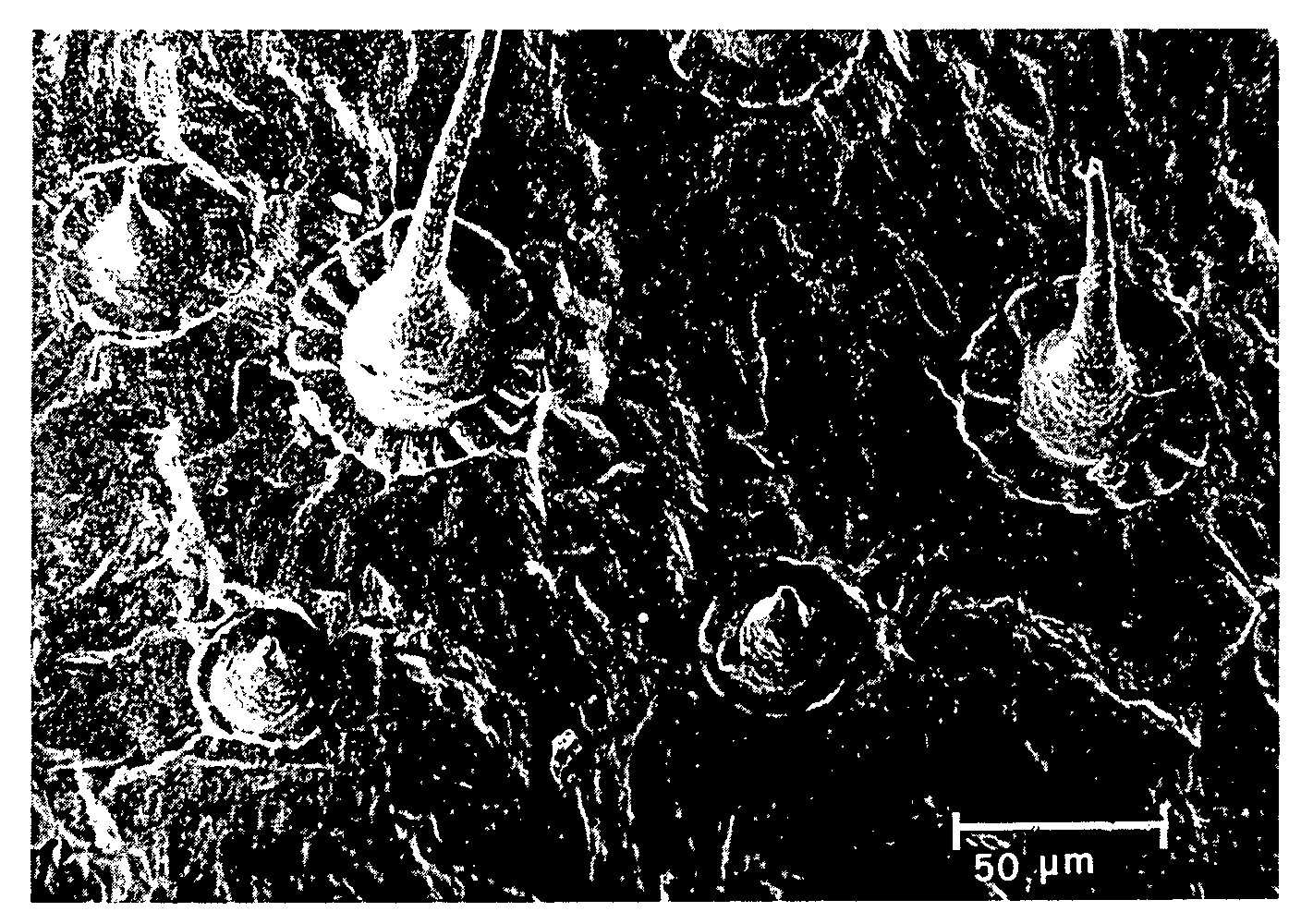

The epidermis of the upper surface of the leaf appears to be composed of polygonal cells covered with a thick cuticle with close-set striae, sometimes in a fan-shaped pattern. It has many cystolith hairs (fig. 6), at various stages of development and more abundant along the veins. These hairs have a verrucose surface and are surrounded at the base by numerous cells (up to 20), arranged in a rosette pattern, more prominent than the surrounding cells, and covered with the cuticle with spoke-like striae. Stomata are infrequent and, in most cases, arranged near the median vein of each leaf segment. Occasional sessile glandular hairs can be detected, also arranged near the veins (fig. 7).
On the lower surface of the leaf (fig. 8), the epidermic cells are smaller than those of the upper epidermis and their cuticle is less thick and briefly striated. There are very numerous stomata and abundant unicellular covering hairs, elongated, conical, with verrucose surfaces, straight or bent, and with a sharp apex. Some hairs are anastomosed with neighbouring hairs. The glandular hairs are numerous, sessile, swollen or collapsed (fig. 8). On the veins, the covering hairs are more abundant, longer and more tapering, pliant and bent; the cystolith hairs are long and slender; glandular hairs are infrequent, and sometimes have short stalks.
The epidermis of the upper surface of the floral bracts is similar to that of the leaves. The cells are polygonal, covered with thick striated cuticular scales. There are fewer cystolith hairs than on the leaves; along the veins they are shorter, whereas in the spaces between the veins they are longer and the cells surrounding their base protrude beyond the neighbouring cells. Near the veins, occasional sessile glandular hairs may be detected.

The lower surface of the floral bracts comprises very numerous stomata, and slender unicellular covering hairs, with a smooth surface, pliant and bent. On the veins, the hairs are more robust and longer, with verrucose surfaces; the base of some is swollen. In the spaces between the veins, there are very numerous glandular hairs, some of which are sessile and others have short stalks; these hairs are fewer in number along the veins and, in the bracts of infructescences at maturity, their stalks are considerably more developed than those of the inflorescences.
The " perigonal" bract, or covering bract - which some authors incorrectly term the tepal bract, covers the perigonium and pistil of the female flower [ [ 1] ]. Most of the resin is produced by its secretory glands, which is why it feels sticky to the touch.
The external, or lower, epidermis of the perigonal bract (fig. 9) is composed of polygonal cells with thick edges and covered with a thin striated cuticle; the infrequent stomata protrude slightly in relation to the other epidermic cells. The few covering hairs are unicellular, rigid, with verrucose surfaces, and bent, particularly along the veins. Cystolith hairs, which are often broken, can also be detected between the veins. Near the edge of the bract, the cells have thinner edges, the stomata are more numerous and the covering hairs are more pliant and more pointed.
There is a very large number of glandular hairs of varying length, some of which are stalked; in some cases their head is swollen with resin and smooth and in others collapsed and devoid of resin. These hairs are more numerous on the veins. In the area between the veins, beside the stalked glands referred to above, there is an approximately equal number of swollen sessile glands, some of which are very large and others considerably smaller (fig. 10). Near the edges the glands are generally sessile and of small size.
On the perigonal bracts which cover the ripe fruit, the stalked glandular hairs have a proportionately longer foot, and in many cases their heads are held together in clusters by the copious resin (fig. 11).
The internal , or upper, epidermis of the perigonal bracts is composed of rectangular cells, with curved and distinctly thick edges and an irregularly striated and thickened surface. Stomata are very infrequent and the unicellular covering hairs, which have smooth surfaces and are flattened, are so compressed by the adherence of the bract to the pistil that they frequently bear the imprint of the epidermic cells themselves (fig. 12). It is also possible to detect slender cylindrical hairs, and near the edge, glandular hairs with short stalks, pear-shaped tips, bent and lodged in hollows in the tissue of the bract itself (fig. 13).
Of the elements observed, those which are of value as regards the production of resin, to which the biological activity of the drug is related, are the glandular formations which, as we have noted, are present in all the aerial parts of Cannabis sativa L. They are particularly numerous on the external surface of the perigonal bract, but also numerous on the lower surface of the leaves and floral bracts, and on the petiole. They are also present, although in very small quantities, on the upper surface of the leaf and the floral bract, on the inner surface of the perigonal bract and on the green parts of the stalk.

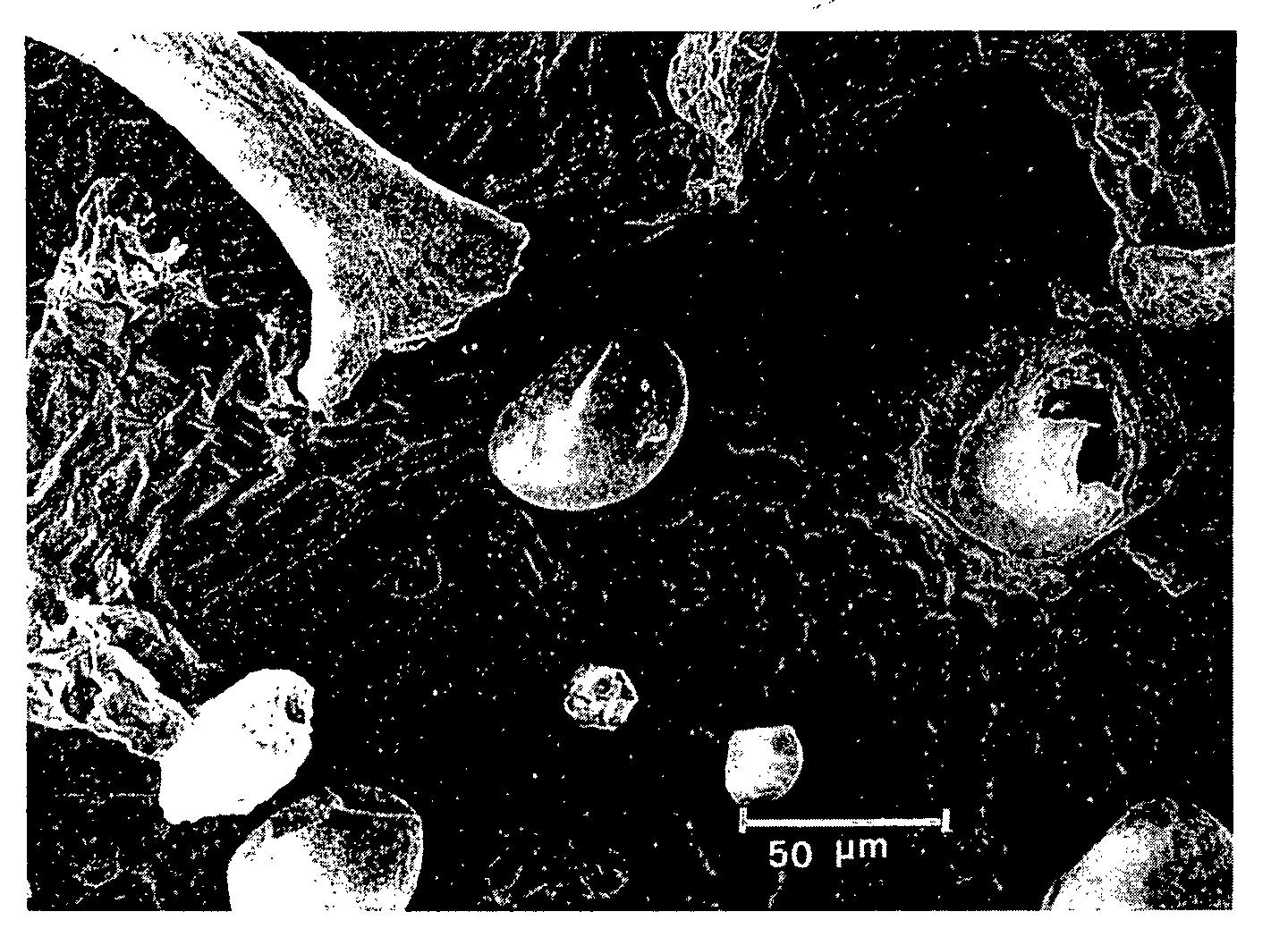




These glands may be divided fundamentally into two types: ( a) stalked or capitate glands and ( b) sessile glands.
The stalked glands were detected in considerable numbers on the external surface of the perigonal bract, and in smaller quantities on the lower surface of the floral bracts; on the other hand, they are very infrequent on the lower surface of the leaves.
They are composed of a stalk, which achieves maximum development on the bracts when the fruit attain maturity. The stalk, which is multicellular and multiseriated, is surmounted by a head, which is more or less swollen, composed of numerous irregular and rather thick polygonal cells with undulating edges.
The stalk, when examined under the transmission electron microscope [1, 3], is seen to be composed of a row of cells of epidermic origin, which cover one or two rows of cells that are similar to those of the palisade tissue and contain photosynthetically active chloroplasts. These glandular formations may nevertheless be regarded as "emergences" since the parenchymatous tissues situated below the epidermis also participate in their formation.
To the two apical cells of the stalk is attached the head, composed of two concentric layers of cells arranged in a rosette pattern and covered with a thin cuticle. The secretion produced by the cytoplasm, on passing through the pores [1, 3], accumulates on the outer edge of the external layer of the head cells and causes the cuticle to become detached by raising it and distending it like a vesicle. The secretion comes out through the openings which are formed by the rupture of the cuticle following the pressure exerted by the secretion or by movements caused by mechanical agencies. As they grow larger, these openings may join in long fissures from which emerges the resin (figs. 14 and 15) which clots on the hairs and in many cases sticks them together (fig. 11).
The " Sessile" glands are present in small quantities on the epidermis of the stalk and branches, in large quantities on the petiole and on the upper surfaces of the leaves and floral bract, and in even greater quantities on the lower surfaces of the leaves and floral bracts. On the external surface of the perigonal bract, they are very numerous - almost as numerous as on that of the stalked glands.
These glands take the form of a hemisphere of variable diameter, with a taut or more or less wrinkled (fig. 16) surface. When detached and observed laterally, they too, however, can be seen to have a short foot.
These glands cannot be regarded as a primary phase of the development of the stalked glands described above, because, unlike in the case of the latter, only the epidermic layer is involved in their formation. In addition, they are full of resin, and when this is discharged, they shrivel and cease to perform their function.
On the external surface of the perigonal bract, particularly near the edges, there are also many glands, but of much smaller size, in which it is possible to distinguish the cells which compose the head, because the cuticle, in most cases, is not dilated by resin. Glands of the same type also exist on these bracts after the development of the ovary into a fruit.
It is interesting to note the presence, on the internal surface of the perigonal bract, of infrequent hairs with short stalks and ovoidal heads, which are lodged in hollows in the surface caused by the adherence of the bract itself to the pistil.
Cannabinoids exist in all the aerial parts of the female plant Cannabis sativa L. These substances may also be observed in the roots, albeit in smaller quantities.
The absence of external glandular formations in the root and the small number of such formations in the stalk lend strength to the hypothesis that, in addition to the external secretory tissues, there are internal secretory cells which contribute to the formation of the resin.


We have described the characteristics, observed under a scanning microscope, of the epidermic tissues of the stalk, petiole, leaves, and floral and perigonal bracts of the female plants of Cannabis sativa L., grown at Messina.
Covering hairs and cystolith hairs are present in all the above-mentioned parts of the plant.
Stalked glandular hairs may be observed in considerable quantities on the lower surface of the floral bracts and on the external surface of the perigonal bracts, and in smaller quantities on the lower surface of the leaves; they do not exist on the upper surface of the leaves and floral bracts, or on the petiole, stalk and branches.
Sessile glands exist in all the parts observed; they are far more abundant on the lower surface of the leaves and floral bracts, and on the external surface of the perigonal bracts, as compared with the upper faces and internal surface of these organs.
The presence of the glandular elements is related to the production of resin in the various parts of the plant.
A. De Pasquale (1970 a). "Farmacognosia della Canape indiana. II. Ricerche morfologiche". Lavori Istituto Farmacognosia Università Messina , 6: 9.
002A. De Pasquale (1970 b). "Prime osservazioni al microscopio ellettronico sulla Canape indiana". Lavori Istituto Farmacognosia Università Messina , 6: 1.
003A. De Pasquale (1973). "Ultrastructure of the Cannabis sativa L . glands". Planta Medica (in the press).
004A. De Pasquale, G. Costa, A. De Sarro (1973). "On the variations in the relationship between some active constituents of Cannabis sativa L . present in different parts of the plant." Paper submitted at the Congress of the Italian Society of Pharmacology, Rome, 4-6 October 1973.
005J. W. Fairbairn (1972). "The trichomes and glands of Cannabis sativa L ." Bulletin on Narcotics, Vol. XXIV, No. 4, page 29.


