A scientific estimation of the patterns of marijuana use
FIGURE
Pharmacological classification (1), (2), (5)
Biochemical actions of marijuana (l), (2), (3), (5)
Effects of marijuana on cell division (1), (2)
Effect of marijuana on spermatogenesis of man (2), (6), (7)
Developmental effects of marijuana (2), (8)
Therapeutic potential
Effect of marijuana on central nervous function (4)
Interaction with other psychoactive drugs
Tolerance
Physical and psychological dependence: drug-seeking behaviour
Long-term effects on the central nervous system (1, 2, 4, 9, 14, 15, 16, 17, 18, 19, 20, 21, 22)
Conclusion
Summary
Author: Gabriel NAHAS
Pages: 13 to 27
Creation Date: 1977/01/01
In this Review, I would like to summarize some of the biomedical findings which have resulted from cannabis research in the past 3 years. In 1976, 3 major monographs (1, 2, 3) dealing with this subject were published; these have to be added to existing comprehensive studies only a year or two older (4, 5). Certainly it is impossible to analyse in any effective manner such monumental work. I will therefore be very selective and retain only the more salient studies which describe some of the basic biological effects of cannabis which may account for some of the clinical manifestations of its acute or chronic usage.
It is essential for estimating the consequence of use of any drug to know how much is taken and how often. For cannabis, it seemed at first as though there were two groups of users: adolescent occasional experimenters, and the wholly different heavy daily user of the Middle East and Far East. A result of this dichotomy was to cast doubts on reports of heavy use in adolescent groups of Western societies. Along with this has gone the view that "drug-users" are in some way a special population, whether from inborn physiology or personality or from social forces - even though no practically successful predictor or proneness to drug use has yet been found. A good deal of light is thrown on all this by the evidence that there is a continuous spectrum of rates of drug use, with no sharp dividing line between experimental, or "moderate", users and heavy users. The evidence, so far as cannabis is concerned, is still entirely derived from answers to questionnaires, and can be analysed in a way similar to that introduced by Ledermann for alcohol consumption. Here a "cumulative distribution'' of rates of use is constructed by calculating the proportion of the population studied who (for instance) used cannabis 100 times or more in the last year, then the proportion Who used it 10 times or more, and so on, recruiting an increasing proportion of the population as successively lower rates of use are used as a reference point. These proportions of the population are then plotted against the rates of use involved. But this is not done directly, but using a probability ("probit") scale for the proportions, and a logarithmic scale for rate of use. The implication of these scales (a "lognormal" plot) is that if there is only one over-all population of individuals varying randomly in rates of use (as they would, for instance, in height), and if the relevant variable in dose is proportional change rather than arithmetical change (as is known for many drugs), then a straight line will result; but if there are two distinct groups, a discontinuity will appear. In the outcome (see figure) there is a remarkably consistent pattern, extending over a considerable range of drugs besides cannabis, and applicable to schoolchildren, undergraduates and soldiers. It should be stressed that such a fit to this particular mathematical pattern is not itself of great significance; it indicates that multiple forces are at work leading to drug use and that the forces at work appear to operate, to greater or lesser degree, in us all. The result does not eliminate the idea of "proneness" to drug use, but suggests that such a quality is not sharply specific to a limited group.
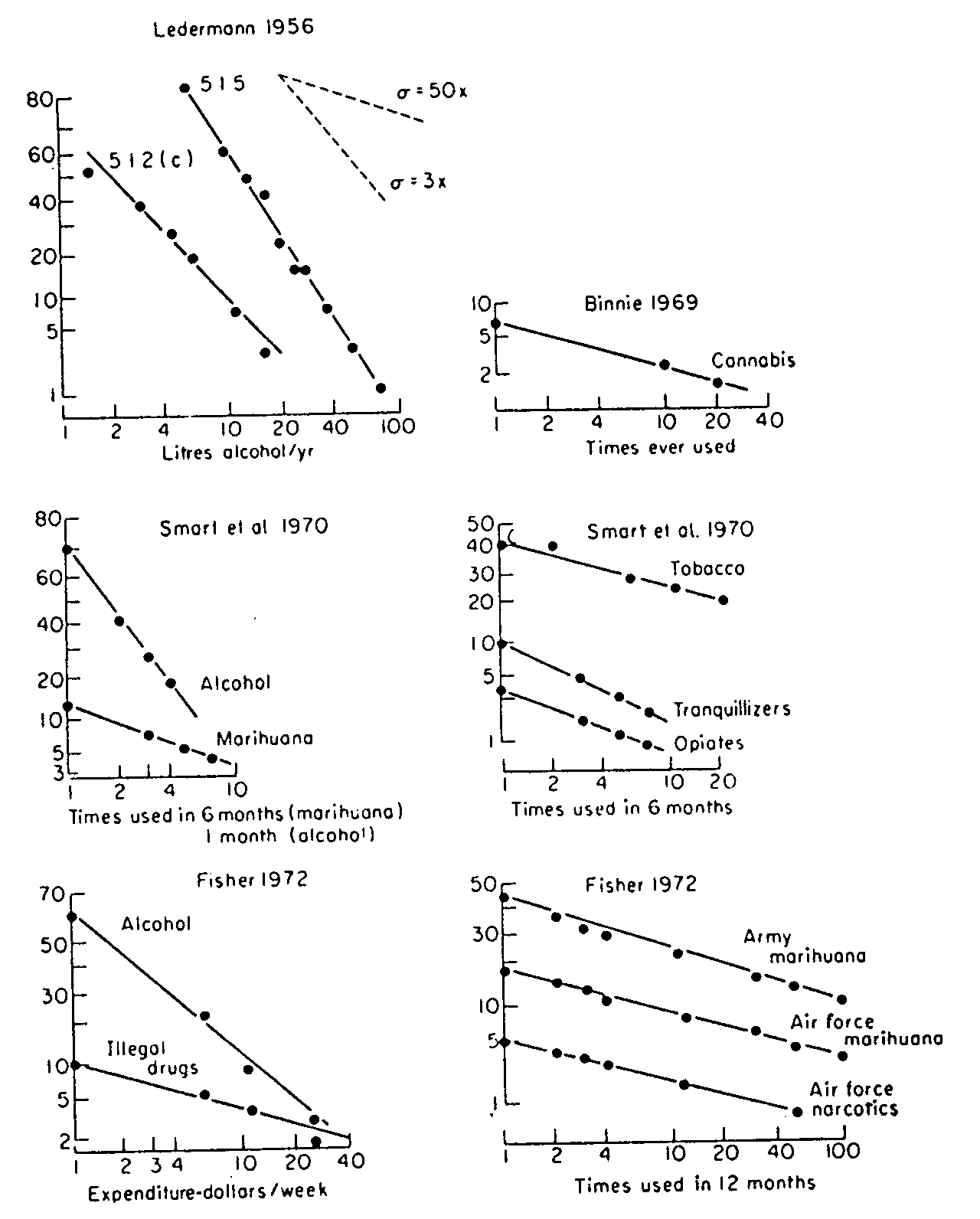
Cumulative distributions of rate of use of various drugs. Horizontal axes, rate of use of drug, on a logarithmic scale; vertical axes, per cent of the population studied who used the drug at a given rate or higher, on a probability scale. The data refer to Canadian schoolchildren, British undergraduates, American armed forces, and French adults and Italian immigrants.
The pattern is similar for cannabis, tobacco, tranquillizers, opiates, "narcotics" and "illegal drugs", but is steeper for alcohol; it appears to be the same for low over-all rates of use of Cannabis as for high over-all rates. (From W. D. M. Paton (1975). Third ISDD Cannabis Symposium.)
For the medical investigator this result provides some clarifications. It allows, in principle, a clearer definition of the magnitude of vulnerable groups, i.e. those with a consumption greater than some limit. It also makes clear the choices confronting those who wish to work towards reducing drug use: namely, either to try to steepen the line by reducing consumption at high rates; or to accept that the general pattern is unalterable, and to reduce the over-all rate of use at all levels (these choices amount to altering the standard deviation or the median of the distribution); or to try for both. The nature of the choice, or indeed the justification for attempts to manipulate at all, is controversial. But it gives some realism to the discussion to appreciate that, for a population of which 20 per cent has tried cannabis at least once in the last 12 months, around 5 per cent will have been using it twice a week or more often.
In a school population, such as the one in San Mateo, California where 55 per cent of the students reported using marihuana during 1975 at least once, it can be projected that 20 per cent of the students used the drug twice a week or more. This would put them medically in a vulnerable position. This projection fits well with the results of a national survey reported by Dr. Robert Dupont, Director of NIDA, which indicates that 8 per cent of the graduating class of 1976 were daily marijuana users and that 53 per cent of the same population used the drug during the same year.
As to the place of cannabis in the family of pharmacological agents, for many purposes the most useful single fact is that of the high fat-solubility of the main active principle (THC) and its first metabolite. For these the octanol/ water partition coefficient is over 1000, and it is only with further metabolism and conjugation that they are brought to forms sufficiently water-soluble to be readily eliminated from the body. From this derives the persistance in the body and the cumulation with repeated dosage. Fat-solubility also confers the ability to penetrate all parts of the body, and radio-active label has been found in every tissue or fluid tested after injection of labelled THC. But the mere presence of a chemical in a tissue is, a priori, neither harmful nor beneficial; and the fact of cumulation merely means that if there are harmful effects, the chronic user will be particularly vulnerable. Fat-solubility does, however, bring a range of pharmacological actions in its wake, linked with those of the anaesthetics and industrial solvents, actions which there is good reason to believe depend on the insertion of the fat-soluble molecule into the lipid of cell membranes. It is only to be expected that such a process will modify cell function, and there is abundant evidence that THC shares this effect with other lipophiles.
Marijuana products have a "half life" of 7 days. This means that after a week, only 50 per cent of the substance is eliminated. Anyone who uses marijuana more than once a week cannot be drug free, and there is a drug build-up in the tissues.
In minute amounts chemicals contained in marijuana (cannabinoids) disrupt cellular metabolism and prevent the proper formation of DNA, RNA and proteins, the building blocks essential for cell division and growth. Scores of scientists have shown that cannabinoids decrease the incorporation of leucine into protein, uridine into RNA, thymidine into DNA, and choline into phospholipids, in brain, testis slice, or cell culture [ (2)] . These effects are evident at concentrations upward of 3 μM, but are occasionally seen at lower concentrations as well. Others have reported uncoupling of oxidative phosphorylation in liver, activation or inhibition of ATPase [ (1)] , [ (2)] (according to conditions in liver, cell culture or red cells); stimulation of adenyl cyclase; inhibition of liver microsomal enzymes; inhibition of prostaglandin synthesis in seminal vesicle, kidney and synaptosomes.
Some scientists disagree as to the mechanism of the interference of THC with synthesis of proteins and nucleic acids (however they agree that the effect is general, rather than limited to a single precursor). Jakubovic and McGeer [ (2)] have not found evidence that uptake of precursor is impaired and believe that a fall in ATP cells and in phosphorylated products might account for the failure of incorporation. Blevins and Regan [ (2)] , who agree that uptake is not impaired, have found a reduction of size of precursor pool that might be due to a leakage of cell membrane, not to a depletion of ATP. Nahas et al. in their studies [ (1)] , [ (2)] of lymphocytes confirm this reduction of precursor pool, but have failed to detect membrane leakage or any depletion of ATP. They believe that the cell membrane is affected in some other way so that uptake is impaired. This difficulty of identifying a single action could be another indication of a general effect of THC that is capable of showing itself in a variety of ways, e.g. on influx, on eflux, or on energy levels.
Many authors have reported that the "cannabinoids'', (chemicals contained in marijuana) whether psychoactive or not, decrease the rate of cell division when added to tissue culture of normal or abnormal cell lines/in 10-6to 10-4M. But more striking is the production of segregation errors during cell division, described by the Leuchtenbergers and Morishima et al. (1, 2) in experiments on tissue cultures exposed to THC in solutions or marijuana smoke. The errors included bridges in anaphase and telophase, unequal segregation, tripolar cell divisions, and the resulting production of "micronuclei", i.e. nuclei containing 30 chromosomes or less against the normal 46. Such in vitro studies could expain observations made in a N.I.H. sponsored study by Stefanis and Issidorides on chronic hashish users, all men, near Athens: 60 per cent showed in their white cells the nuclear drumstick: this is accessory chromatin typical of female cells and normally occurring in less than 1 in 500 men. The observation prompted Stefanis to some histochemical observations on the chromatin of leucocytes and spermatozoa in these subjects. This revealed a reduction in arginine-histone in lymphocytes, and in the arginine-specific staining expected from protamine in spermatozoal heads.
All of these cellular effects of marijuana in concentrations which may be reached in human consumption raised the possibility that chronic use of marijuana might affect spermatogenesis and impair reproductive function.
The impairment of spermatogenesis in man by marihuana smoking has been best illustrated by an interdisciplinary study I carried out with Dr. Hembree, Morishima, and Zeidenberg at the College of Physicians and Surgeons and the Psychiatric Institute of Columbia University.
Over a 2-year period 16 marijuana smokers aged 18 to 23 were hospitalized in the Research Ward of Psychiatric Institute of Columbia University. Each subject was studied for 8 to 12 weeks. The study was divided into 3 periods: a 3- to 4-week drug-free period, a 4-week smoking period, and a 3- to 4-week wash-out phase. All subjects were carefully selected from a large pool of marijuana smokers, and only those in good physical and mental health were retained. During the smoking period, the subject smoked an average of 5 to 15 cigarettes of marijuana a day (0.9 gm of marijuana, 2 per cent THC). The highest daily consumption was 36 in one subject. The subjects claimed to have smoked such large amounts on certain occasions, when marijuana was available.
Twelve of the sixteen subjects had significant and sustained decrease in sperm concentration occurring only after 2 weeks of marijuana smoking and sustained for at least 2 weeks. All counts were normalized to 200, based upon the average obtained during the drug-free control period. Group sperm counts decreased significantly only during the first and second week following the end of smoking. The average decrease for all 16 subjects was 35 per cent with individual decreases ranging from 30 to 72 per cent. Decrease in sperm count was associated with a decrease in spontaneous motility which was most marked immediately after the end of smoking. Statistical significance was maintained for only 2 weeks because of the large variance.
Finally there was also a significant decrease in normal forms which was noted at the end of the smoking period and sustained for an additional 3 weeks. All of these changes took place in the absence of any significant changes in the levels of testosterone, L.H. and F.S.H. The time course of the changes is consistent with an effect on spermiogenesis. During this period, gene activity gradually decrease as the permatid chromatin is packaged within the dense matrix of the newly synthesized arginine-rich protamine. Interference with transcription, translation, or both, resulting in failure to synthesize the structural proteins required for axonemal development and the formation of the other specialized morphological elements of the mature spermatozoon, is likely to be associated with decreased sperm in the ejaculate 12-45 days later. Therefore, if it can be assumed that the decreased count noted reflected a specific perturbation of germ cell production 28 days earlier altered spermiogenesis induced directly by the cannabinoids best explains the observations.
The most potentially damaging effect of marijuana on spermatogenesis is the marked increase in abnormal forms of sperm cells. This raises the possibility of a genetically transmitted abnormality if a viable sperm with decreased genetic information fertilizes an egg.
Such possibility could only be resolved by epidemiological studies performed on children born to marijuana smoking parents.
In the meanwhile, there is evidence that marihuana produces such abnormality. This evidence is conflicting when one considers all of the teratogenic studies performed on rodents which have a short gestation period. Some studies report increase in birth defects in the treated animals, others do not. However, a study on rhesus monkeys by Sassenrath and Chapman indicates that delta-9 THC chronically administered produces impairment in reproductive function. The following is an abstract of their work:
"The reproductive success of long-term chronically THC-treated rhesus monkeys has been compared to that of undrugged cagemate controls. The data presented are from 27 matings over three successive birthing seasons, involving 5 THC-treated and 5 undrugged females paired with 2 THC-treated and 2 undrugged males in various combinations of drugged and undrugged males and females.
"In this study, drug exposure of young adult breeders was continual and daily over the three-year period. THC was given orally on preferred food at 2.4 mg per kg once daily for the first two years and was increased to 2.4 mg per kg per day given twice daily prior to the third breeding season. All chronically drugged subjects were behaviorally tolerant to the 2.4 mg/kg/day dose and showed only mild sedation effects on the double drugging.
"Although over-all conception for all breeder pairs involving at least one THC-treated parent was not appreciably different from controls (94 per cent compared to 100 per cent), the over-all reproductive success (taking into account foetal wastage, stillbirths, neo-natal deaths and infant deaths) was markedly lower for drugged parents (59 per cent compared to 90 per cent). Histopathologies in non-viable offspring were observed only in those with at least one drugged parent, and included hydrocephalus, atrophic thymus, myocardial and umbilical degeneration, inguinal and umbilical hernias, and abnormal pancreas (atrophic and ectopic). Many or all of these have been reported in rodent offspring of mothers treated with THC or cannabis at higher dose levels, and are observed rarely in routine autopsy of infant deaths in the total rhesus breeding colony.
"Although the sample size is small, the data suggest that:
Failure to conceive or resorptions are associated with THC-treatment of the female parent;
Abortions, stillbirths, and neo-natal deaths are associated with THC-treatment of the male parent;
Reproductive deficits do not tend to become more frequent or severe over successive breeding seasons for the same breeder pairs as long as treatment conditions remain constant. However, abortions and resorptions were observed only during the third birthing season after the daily THC dosage has been doubled.
"The effects associated with THC-treatment of males is of particular interest and raises the question of possible chromosomal drug effects via the sperm. Behavioural and endocrine data on these breeding groups do not strongly support the stress hypothesis." The same authors observed that 6 male surviving offspring presented heightening of locomotor activity and increased behavioural responsiveness to visual and auditory stimuli.
Three actions should be mentioned for possible therapeutic use: first, bronchodilation. Inhalation of delta-9-THC causes a considerable increase in airway conductance; it is not antagonized by a -blocker, and so is not due to sympathetic activation. It does not prevent mechacholine bronchoconstriction, so it is not an atropine-like action. The effect is most probably due to the ability of anaesthetics to relax smooth muscle. The therapeutic use of THC inhaled as a smoke may be limited. Repeated marijuana inhalation in fact diminishes airway conductance and maximum expiratory function, even though an inhalation temporarily restores the normal situation. Here is a novel approach to bronchodilation, however, which ought to be detached from its cannabinoid environment and examined in its own right. The main problems are, delivery of the drug and the side-effects.
The second possible therapeutic use is based on the ability of the cannabinoids to lower intraocular pressure by up to 30 per cent. This can be demonstrated in patients with glaucoma as well as in normal subjects. This effect is not due to a general lowering of blood pressure. In a study using delta-9-THC and two psychically active metabolites, cannabinol and cannabidiol, the order of effectiveness in reducing intraocular pressure (IOP) was the same as the order of effectivenss in producing a "high". This lends support to the idea that it is a central effect, perhaps related to sedation. It has also been reported that THC and cannabinol are active by direct application to the eye. Further, THC, which with other cannabinoids is known to inhibit prostaglandin synthesis at about 10 - 100 μM, was able to reduce the rise in IOP produced by arachidonic acid infusion.
The third potential therapeutic effect of marijuana is in the treatment of convulsive seizure. It has been shown in a number of animal preparations that cannabidiol (not THC) compares well to phenobarbitol and Phenylhydantoin as an anticonvulsant. Because of the very low psychoactive effect of cannabidiol, it would appear that it may be a useful antiepileptic agent. Its mechanism of action remains to be determined but it might be related to its depressing action on neuronal activity.
If marijuana products are proven to be useful therapeutic agents, their usefulness might well be attenuated if marihuana is widely used as a "recreational drug".
Out of hundreds of experimental papers on animals, it is now established that marijuana products affect all parts of the brain, but that the primary physiological and chemical changes produced by this drug occur in the so-called limbic diencephalic structures or paleocortex (old brain).
Penfield and Jasper showed in 1954 ( [ 10)] that abnormal discharges in or near the limbic cortex may produce feelings of depersonalization, distortions of perception, alterations in time sense, and feelings of fear or paranoia. All these subjective states may occur and some are very common as part of the marijuana experience. Pleasant feelings, euphoria, happiness and placidity are also very common elements, and the septal region which appears to function as a major coordinating centre for the entire limbic system, is by far the most effective target for self-stimulation experiments with a variety of mammals, including man. Delgado [ (11)] has demonstrated that electrical stimulation of limbic structures, especially the hippocampus, often produces pleasant sensations, elation, deep thoughtful concentration, relaxation, and coloured visions in human subjects with chronically implanted depth electrodes.
Studies by Heath [ (12)] and by McIsaac et al. [ (13)] provide objective evidence that the primary physiological and chemical changes induced by marijuana do occur in limbic-diencephalic structures. Heath recorded electrical activity from multiple subcortical brain regions and from scalp electrodes in one psychiatric patient. Recordings were made repeatedly over several weeks during all states of consciousness and during intoxication with marijuana, alcohol, and amphetamines and while smoking tobacco. Only during marijuana intoxication and only associated with "rushes" of euphoria, Heath recorded marked changes in electrical activity patterns from the septal region. There were no significant changes in the activity in any other area including the scalp-recorded EEG.
McIsaac et al. injected squirrel monkeys with radio-active delta-9-THC and, using radioautographic techniques, showed that very high concentrations appeared in the limbic system, diencephalon, mid-brain, and frontal and cerebellar cortex within 15 minutes and remained in these regions at higher concentrations than in other brain areas for up to 4 hours. They also noted a different effect of dose on the behaviour of the monkeys, with low doses producing apparently diminished anxiety, moderate doses inducing stimulation, and high doses producing incapacitation.
This may be the most inclusive neural model of the physiological basis of the marijuana experience: delta-9-THC and its metabolites act primarily to alter the normal functional relationship between paleocortical limbic system structures ("old brain") and the neocortex ("new brain"). This alteration may vary from stimulation of limbic activity to depression (or disinhibition) to increasing inhibition), depending on dose, time, previous experience and current mood. The major elements of the marijuana experience, including altered perception, mood and performance, may all be explained on this basis. The most striking objective change, i.e., a general cognitive performance decrement, may be the result of the loss of an accurate concept formation, appraisal, or evaluation stage in the stimulus-cue-performance sequence, normally subserved by neocortical limbic circuits. The occasional "bad trip", which has been reported in experimental subjects, seems to occur when the individual is already feeling badly or is apprehensive about the experimental situation, and emphasizes the importance of past and present experience in determining the quality of the emotional aspects of marijuana intoxication.
The model would explain not only the major elements of the acute marijuana experience itself but also the occasionally reported "flashback" phenomenon and possibly some of the cannabis-mobilized psychoses. It is well known to neurophysiologists that the limbic system and the hippocampus in particular, has a very low threshold for activation by mechanical, chemical, or electrical stimulation. Once activated, neuronal discharges tend to spread throughout all limbic circuits without very readily involving other brain areas. These phylogenetically old neural structures also have a marked tendency to persist in an altered functional state for long periods after the initial stimulus has been withdrawn. In this regard, it is perhaps significant that Heath has recorded bursts of high voltage spike and slow activity from limbic areas as the only consistent electrographic abnormality in a large number of patients during periods of psychotic behaviour.
The exact cellular mechanisms of these multiple changes in the central nervous system are being investigated. It is difficult at the present time to combine the reports on the roles of Acetylcholine, catecholamines, 5-HT, and prostaglandins into a coherent story. They have all been implicated by different investigators.
It might be useful to think of THC as lifting inhibitory gates involved in sensory processing: from this could follow logically the sensory effects, the interference with short-term memory, the prolongation of "felt" time against "clock" time, the neurophysiological evidence of hypersynchronous high voltage bursts at various sites, and a number of "release" phenomena-myoclonic jerks and the like. In one recent report, bringing in cerebellar activity explicity, Fernandez-Guardiola et al. (1) have shown that, in the anesthetized cat, THC produces a great acceleration of multi-unit activity in neurones of the sensory cortex, and in the Purkinje cells of the cerebellum, with a slowing of multi-unit activity in the red nucleus (believed to be under inhibitory control of Purkinje cells).
One should not discount the possibility that THC and its metabolites might also impair in the central nervous system the uptake of precursors of the proteins (enzymes) which elaborate the neurotransmitters which are continuously synthetized within neurone. It would be most important to study the effects of the cannabinoids on the fast and slow axonal flow of these precursors along the neurone.
This is an immense subject which has been well summarized by the behavioural study of Pryor (1976) who researched the interaction of delta-9-THC with 13 other drugs (2). A battery of tests, 13 largely behavioural, 3 physiological, were used: the results for each drug alone and combined with THC were pooled in two ways, first expressed in terms of the standard deviation of the observations so that their magnitudes are related to the size of normal variation in response; and second, regardless of sign as between "stimulation" and "depression", or taking account of this. A major acute interaction of delta-9-THC with other drugs is a mutual potentiation of any depressant properties, and an antagonism of any stimulant properties. The principal conclusion, borne out by all this work is that when THC is added to any of these drugs, the resultant effect either shifts to the depressant side, or becomes more depressant. This indicates something about mechanism of action because it is similar to the way that anaesthetics reduce the excitant effects of neuromuscular blocking agents and increase their blocking action, and to Paton's theory that THC is related to general anaesthetics.
All euphorigenic or sedative drugs, if given for some time, evoke an adaptive response, and-on withdrawal of drug-some sort of rebound: the only question is, what are the characteristics of response and rebound?
In studies performed at Psychiatric Institute of the Columbia-Presbyterian Medical Center, marijuana cigarettes were made freely available to hospitalized volunteer subjects. These subjects smoked an average of 10 to 20 marijuana cigarettes (.9 gm cigarettes, 2 per cent THC) daily for a period of 4 weeks. This is an amount comparable to that used daily by chronic cannabis smokers in Jamaica, Costa Rica, or Morocco. All presently available pharmacological and clinical evidence indicates that frequent (daily) users of cannabis develop tolerance to the physiological as well as the psychological effects of the drug. This tolerance to cannabis gives a physiological basis to the necessity for the frequent smoker to increase dosage or to seek more potent psychotropic drugs such as other hallucinogens or the opiates. However, it would appear that this tolerance to the effects of marijuana does not involve the effects of this drug on the triggering of abnormal activity in the limbic system, according to the work of Heath. Neither is there a tolerant effect on spermatogenesis since all chronic marijuana users that we observed had an increased incidence of abnormal forms of sperm and a lowered sperm count.
A misconception seems to have penetrated into the minds of many psychologists and physicians that "addiction", meaning physical dependence accompanied by withdrawal symptoms, is the main criterion by which the potential harm of a drug to the individual or to society should be gauged. There is a very fine line between physical and psychological dependence. There is no complete dichotomy between mind and body. Psychological function also has physiological and biochemical bases. The desire for instant gratification is a profound psychological reinforcer. Physical dependence does not develop with central nervous system stimulants such as cocaine, which is known to create in an individual one of the most enslaving types of drug dependence. Addiction to a drug is not a function of the ability of the drug to produce withdrawal symptoms. Drug dependence results basically from the reproducible interaction between an individual and a pleasure-inducing biologically active molecule. This interaction leads to what Wickler calls drug-seeking behaviour or behaviouraldependence (5). The common denominator of all drug dependence is the psychological reinforcement resulting from reward associated with past individual drug interaction, and the subsequent increasing desire for repeated reinforcement. On this basis, it is deceptive to categorize marijuana as a "soft" acceptable drug which does not create dependence.
Although cannabis users develop tolerance to the drug, they do not present any significant physical dependence identifiable by specific withdrawal symptoms similar to those occurring with heroin or ethanol. The symptoms observed following discontinuation of heavy use are relatively mild. Loss of appetite, insomnia, and irritability are well tolerated, but it is well documented that cannabis may create a state of psychological dependence or behavioural dependence which is an important obstacle to discontinued usage.
In man, physiologic marihuana abstinence signs have not been demonstrated, but behavioural (and some physiologic) abstinence phenomena have been reported in heavy users of hashish or ganja. The between-dose hyperirritability and dysphoria reported to occur in experimental studies on chronic marijuana intoxication may actually be early and short-lived abstinence changes (14).
In the West, where marijuana with relatively low delta-9-THC content is widely smoked, dependence in the sense of drug-seeking behaviour appears to be less a function of any pharmacologic reinforcing properties the drug may have than of secondary (conditioned) reinforcement derived from the social milieu in which the marijuana is smoked. In cultures where marijuana of higher delta-9-THC content, hashish, or ganja is used, pharmacologic reinforcement (through suppression of abstinence changes) may play a greater role in maintaining drug-seeking behaviour.
This is an area of great controversy mainly because there is no satisfactory method to assess in man damage to the limbic system, or to appraise changes in personality. Some investigators [ (13)] who have studied groups of chronic cannabis users in Jamaica, Costa Rica and the US have concluded that there is no amotivational syndrome and no evidence of cerebral dysfunction of psychosis. On the other hand, investigators from Egypt [ (16)] , India [ (17)] and Jamaica [ (18)] have pointed out the existence of such effects in groups of chronic cannabis users they have studied. In the United States, opinion is sharply divided as well: Fink [ (15)] , Mendelson (19), Freedman [ (15)] , claim that cannabis use is not associated with serious mental pathology, whereas Kolansky and Moore [ (20)] , Hart [ (21)] , and Powelson have concluded just the reverse. Only time and the performance of carefully controlled longitudinal studies will resolve this controversy.
Meanwhile, the well-controlled animal experiments that have already been performed should be carefully appraised. These have been done by Heath [ (1)] , [ (2)] , [ (22)] , by Sassenrath and Golub [ (9)] , and by Kalant [ (2)] . These studies all point to a persistent damaging effect on the mammalian brain of chronic cannabis usage in dosages reached in human consumption. A summary of these three studies follows.
The following résumé of the experiments of Heath have been sent specifically for inclusion in this report:
"We have found in our studies of marijuana in rhesus monkeys that exposure to active marijuana smoke and to delta-9-THC at a frequency of five times per week induced changes in the behaviour of the animals and, more importantly, in electroencephalographic (EEG) recordings from precise deep regions of the brain. Brain regions affected were those sites where activity has been correlated with emotional behaviour. The EEG changes were of the kind that could account for the reduction in motivation and drive and the general apathy that one sees in chronic users of marijuana. Changes were not apparent on conventional scalp EEGs.
"When the monkeys had been exposed to the cannabis sativa derivatives for two to three months, lasting alterations in brain function occurred. After 6 months' exposure, we continued to follow the animals for an additional three-month period, and the recording abnormalities persisted. That early work has been reported.
"Our studies were carefully controlled. Some monkeys in the group were exposed to inert or inactivated marijuana on the same smoking schedule as those animals that were heavy smokers of active marijuana. None of the control monkeys showed behavioural or EEG changes. Other monkeys were given delta-9-THC intravenously and they showed the same changes in brain function as the animals that were heavy smokers of active marijuana. It was therefore evident that it was the active ingredient that was responsible for lasting brain changes.
"The criticism of our work has not been based on objective findings. Specifically, we have been critized for giving the monkeys excessive doses of marijuana. This issue was discussed with Dr. Julius Axelrod, and he agreed that our dosages were comparable to those used by many human smokers of marijuana [ (2)] . Further, those criticisms did not take into account the fact that monkeys are inefficient "smokers". In contrast to the human, the monkey tends to stop inhaling when exposed to smoke. In our recent studies, therefore, we have developed a method of smoking the monkeys more efficiently. A respirator was modified and a smoking procedure was established to simulate the pattern of human smokers. With this new method, we have been able to use smaller quantities of the active material.
"Some monkeys currently under study are now being exposed to one joint per day, five days per week. The active material in a joint is 0.25 grammes. This is measured on a per weight basis and does not exceed the amount consumed by the average human marijuana smoker. With the new smoking method, simulating the human smoker, the monkeys are having a profound response, both in terms of behaviour and EEG changes.
"We have also carried out some histopathologic studies on the brains of monkeys which were included in the first study. Extensive electron microscopic studies were done on precise brain regions of (1) monkeys exposed to active marijuana smoke, (2) monkeys exposed to inactive marijuana smoke (smoked at the same frequency and at the same dose level), (3) monkeys given delta-9-THC intravenously, and (4) "clean" monkeys which had not previously participated in any study. Brains of monkeys which received active material in the form of smoke or by intravenous injection showed distinct changes in the synaptic cleft and in those regions - septal region, hippocampus, and amygdala - where EEG changes has occurred. Changes were (1) a widening of the cleft, (2) deposition of a dark opaque material in the cleft, and (3) a beginning clefting of the synaptic vesicles at the terminus of the axone. These changes all suggest incipient damage to the neurones. This electron microscopic study has been submitted to the Journal of Neuroscience Research and should be published sometime in the summer."
One might add that the monkeys treated with marijuana kept on eating, sleeping and performing their daily gymnastics.
A preliminary report by E.N. Sassenrath and Golub (9), further indicates that monkeys born to THC-treated parents present alterations in locomotor activity and responsiveness, which could well indicate an alteration of the of their brain., This report reads as follows:
"As a result of a three-year breeding programme using long-term THC-treated monkeys and controls, six infants have been born, one or both of whose parents had received daily THC for up to a year prior to conception. These infants have been tested along with control offspring matched for age, sex, and social experience.
"Two male offspring of THC-treated mothers born in 1974 and 1975 were tested after weaning (at 6 months) for social behavioural adaptation with control peers and for "functional intelligence" (i.e. retrieval of a desired object in a naturalistic problem situation). Both THC males scored significantly higher than four control cagemates on active affiliation (i.e. play and mounting) and significantly lower on passive affiliation (i.e. groom, huddle, and proximal spacing), although they showed no tendency to be more aggressive or more dominant than control peers. In the functional intelligence tests, the THC males solved more problems faster, with higher scores on approach and contract orientation to new problems.
"Four THC-offspring (two males, two females) have recently been tested prior to weaning at 3.5 months of age for behavioural and autonomic responsiveness to a brief separation from mother and exposure to a novel test environment. Initial analysis of the data indicates the offspring of THC-treated parents scored higher on measures of locomotor activity and cardiac and behavioural responsiveness to visual and auditory stimuli.
"These studies are continuing and will be augmented by observations of offspring from the current breeding season. However, the consistency of observations to date strongly suggests a non-specific heightening of locomotor activity and responsiveness to environmental stimuli in offspring of THC-treated parents." (This resembles the hyperkinetic syndrome of the growing child.)
Kehr and Kalant have also reported permanent learning impairment after chronic heavy exposure to cannabis in the rat (2). The dose of THC used orally for six months was 10 mg/kg which corresponds to heavy human use similar to that smoked in our studies at Columbia. The animals were visibly intoxicated for only 4 h after each dose, gained weight normally and were in good health throughout the experiments. After 6 months of treatment there was permanent impairment of learning on 2 different tasks in a food-motivated maze task. The authors state that "EEG changes in the present work tend to support the interpretation that we are dealing with organic damage for which histological confirmation is being sought".
As mentioned before, the extent at which chronic marijuana use will produce in man serious, permanent psychiatric illness such as psychosis, is still a matter of heated debate. However, most psychiatrists, at least at the Psychiatric Institute in New York, believe that marijuana should not be used by any person prone to mental illness such as schizophrenia which this drug might trigger or worsen.
The biologically active molecules contained in marijuana interact with the basic components and processes of living cells. As a result of such interactions, experimental evidence indicates that cannabis interferes with cell division and metabolism, reproduction, embryological development, learning, and growth.
Only longitudinal epidemiological studies of marijuana-smoking populations may document in man the pathologic effects of long-term cannabis usage. To my knowledge the literature does not contain a single autopsy report on a long-term chronic marijuana smoker. Therefore the human pathology of marijuana cannot be written before two or three decades. (It took sixty years for investigators to establish the pathology of tobacco smoking.) Meanwhile, on the basis of their present short-term observations and past experience with other drugs, biologists and physicians can only make certain predictions about what this pathology might be. I would venture that such pathology might develop in two areas:
As a result of the specific disrupting effect of cannabis on the diencephalic limbic area, one might expect an increased incidence in mental illness among cannabis users. Such incidence has been reported by some [ (18)] , [ (20)] , [ (21)] .
As a result of its inhibitory effect on dividing cells, one might expect an increase in the incidence of developmental defects in the offspring of chronic users. These have not been heretofore reported.
A scientific estimation of patterns of use of marijuana indicates that prevalence of use in a given population is obligatorily associated with a high incidence of use in a fraction of this population. The more widespread the usage, the greater the fraction of heavy users. Marijuana use has followed such a pattern in the US: 8 per cent of the 1976 high school graduating class are daily marijuana smokers, while 53 per cent of the same population used the drug during the same year.
Marijuana products (cannabinoids) like THC are fat soluble substances which remain in the body for at least 8 days after a single administration. Anyone who uses marijuana more than once a week can not be drug free.
In minute amounts, cannabinoids disrupt cellular metabolism, and prevent the formation of DNA, RNA and proteins, chemicals essential for proper cell division and growth.
Cannabinoids, whether psychoactive or not, decrease the rate of cell division when added in minute amounts to tissue culture of normal or abnormal (cancerous) cell lines. This decrease in cell division is associated with an increase in the number of abnormal cells which do not contain their proper amount of DNA, the chemical which carries the genetic code. Abnormal white blood cells and sperm cells have been sampled from chronic hashish users.
In a controlled study, 16 young men (in good mental and physical health) smoked 5 to 15 marijuana cigarettes daily for one month. After this time, they presented a decrease in sperm count, a decrease in motility of sperm, and a marked increase in abnormal forms of sperm cells. The possibility of a genetically transmitted abnormality as a result of daily marihuana usage is raised as a result of these observations.
The possibility of genetic damage is illustrated by a study on rhesus monkeys fed THC over a period of 3 years. Failure to conceive or resorptions were associated with THC treatment of the female parents; abortions, stillbirths and neo-natal deaths were associated with THC treatment of male parents (raising the possibility of a drug effect transmitted via the sperm). Six male surviving offspring had abnormal locomotor activity and increased behavioural response to stimuli.
THC might be useful in the treatment of asthma, glaucoma. Another cannabinoid, cannabidiol might be useful in the treatment of epilepsy.
THC acts on the septal area of the limbic system of the brain ("old brain") where structures controlling emotional behaviour are located.
Monkeys who had deep electrodes implanted in the "limbic area" of their brains were studied for 6 months while they were exposed daily to marijuana smoke. Abnormal brain wave patterns persisted 3 months after smoking was stopped. Microscopic studies of the brains of these monkeys showed lesions of the nerve cells in this septal limbic area which controls emotional behaviour.
Many psychiatrists believe that marijuana should not be used by any person prone to mental illness or who has been treated for such an illness which this drug might trigger or worsen.
THC interacts with many other psychoactive drugs either by increasing their depressive properties or by decreasing their stimulant ones.
Daily users of marijuana develop a tolerance to the physiological and psychological effects of this drug.
Marijuana users, when they stop using the drug, do not present withdrawal symptoms similar to those occurring with opiates. However, with absti nence, changes in behaviour and mood have been reported. Daily use of marijuana is associated with behavioural dependence and drug-seeking behaviour.
Braude, M. and S. Szara, Pharmacology of marihuana, Raven Press, New York, 1-838, 1976.
002Nahas, G. and W.D.M. Paton, Marihuana: Chemistry, biochemistry, and cellular effects, Springer Verlag, New York, 1-552, 1976.
003Graham, J.D.P., Cannabis and health, Academic Press, New York, 1-447, 1976.
004Miller, L., Marijuana, effects on human behavior, Academic Press, 1-397, 1974.
005Mechoulam, R., Marihuana: Chemistry, pharmacology, metabolism, clinical effects, Academic Press, 1-409, 1973.
006Hembree, W., P. Zeidenberg and G. Nahas, Oligospermie des fumeurs de chanvre, Bull.Acad. Nat. Med., 160, 102, 1976.
007Hembree, W., J. Huang and G. Nahas, Effects of heavy marihuana smoking on gonadal function of man (in press).
008Sassenrath, E.M. and D.F. Chapman, Long term effects of Δ 9THC on reproduction of the rhesus monkey (in press).
009Sassenrath, E.M. and M.S. Golub, Behavioral characterization of offspring of THC treated rhesus breeders (in press).
010Penfied, W. and H.H. Jasper, Epilepsy and the functional anatomy of the human brain, Little Brown, Boston, 1-386, 1954.
011Delgado, J. M.P., Physiological correlates of emotion, Academic Press, 1-287, 1970.
012Heath, R.G., Brain function and behavior. Emotion and sensory phenomena in psychotic patients and experimental animals, J. Nerv.Ment. Dis., 100, 159, 1975.
013McIsaac, W.M. and J.E. Fritchie, Distribution of marihuana in monkey brain and concomitant behavioral effects, Nature, 230, 593, 1971.
014Wickler, A., Aspects of tolerance to and dependance on cannabis, Ann: N.Y. Acad. Sci., 282, 1976.
015Dornbush, R.L., A.M. Freedman and M. Fink, Chronic cannabis use, Ann. N.Y. Acad. Sci., 282, 1976.
016Souief, M.I., Chronic cannabis users; further analysis of objective test results, Bulletin on Narcotics, XXVII: 1, 1975.
017Chopra, G.S., Psychoclinical effects of long term marihuana uses in 275 Indian chronic users, Ann. N.Y. Acad. Sci., 282, 1976.
018Knight, F., Role of cannabis in psychiatric disturbance, Ann. N.Y. Acad. Sci., 282, 1976.
019Mendelson, S.H., The use of marihuana: A psychological and physiological inquiry, Plenum, New York, 1-202, 1974.
020Kolansky, H. and W.T. Moore, Effects of marihuana on adolescent and young adults, J. Am. Med. Assoc., 216-486, 1971.
021Hart, R.H., A psychiatric classification of cannabis intoxication, J. Am. Acad. of Psych. and Neurol., 1, 83, 1976.
022Harper, J.W., R.C. Heath and W.A. Myers, Cannabis sativa: Its effects on ultrastructure of the synapse in monkey brain, J. of Neurosciences (in press).


