MORPHINE
Author: Harry Peterson
Pages: 23 to 29
Creation Date: 1955/01/01
An accurate knowledge of the transformations of a drug that occur after its ingestion may determine whether it can be chemically detected or not in the body after death or in the excretions or secretions of the living body. It may appear unchanged or as entirely different substances, and the methods of analysis employed must be modified accordingly.
Narcotics constitute a group of drugs of interest both to racing chemists and to toxicologists. Before coming to a discussion of the particular narcotics named in the title of this paper, the general reactions of drugs in the body may be considered.
Chemical Reactions Occurring in the Body
Reactions which can take place in the body include oxidation, reduction, hydrolysis, and a number of synthetic reactions or conjugation processes. R. Tecwyn Williams, in his book Detoxication Mechanisms[1] , lists nine such synthetic reactions, five of which have been reported to occur with the drugs of most interest to racing chemists. They are:
Glucuronic acid conjugation
Ethereal sulfate conjugation
Glycine conjugation
Acetylation
Methylation
Detoxication may involve only one of these reactions, or several may take place successively or concomitantly, depending on the structure of the substance ingested. In the subsequent discussion, most of the material has been taken from Williams (1). Other sources of information are noted as they occur.
Hydrolysis
This reaction occurs with esters like atropine, cocaine, and procaine. The de-acetylation of diacetylmorphine likewise is hydrolytic. In every case the presence of enzymes is important.
Oxidation
Oxidation occurs in a number of ways and is largely dependent on the structure of the compound. Amines, notably many sympathomimetic amines, are oxidized in the body, with enzymes like amine catalysing the reactions. Methyl groups may be removed by oxidation also. In radioactive-tracer experiments with rats given codeine, in which the methoxyl group is labelled with carbon 14, C 140 2 is found in the expired air. Primary alcohols are oxidized to the corresponding acids.
Reduction
Reduction of foreign substances in the body appears to be less common, but a well-known example of a reduction process after the administration of a drug is the reduction of chloral hydrate in vivo to trichloroethyl alcohol. This is usually followed by combination with glucuronic acid to form the trichloroethylglucuronide, urochloralic acid.
CCl 3CHO + 2H CCl 3CH 2OH
Glucuronic Acid conjugation
CCl 3CH 2OH CCl 3CH 2OC 6H 9O 6
Synthetic reactions
Glucuronides

The formation of conjugates with glucuronic acid is probably the most common synthetic process in the body. Two kinds of such glucuronides are known. One is an ether type which is formed by reaction with hydroxyl groups in almost any type of compound except carbohydrates. The other is an ester type formed by reaction with carboxyl groups attached to aromatic nuclei. The ether type is usually stable in alkali and does not reduce alkaline copper reagents, whereas the ester glucuronides are unstable in alkali, and the glucuronic acid liberated reduces such reagents. In the case of urochloralic acid, however, its ability to reduce Fehling's solution is attributed to the instability in the presence of strong alkali of three halogen atoms attached to the same carbon.
The morphine molecule contains both a phenolic and an alcoholic hydroxyl group. In codeine the phenolic hydroxyl is methylated, and only the alcoholic one is for conjugation. Glucuronides are formed with both compounds [2] . Morphine, however, is capable of still another reaction which results in the formation of ethereal sulfates, found together with glucuronides in the urine after morphine ingestion.
Ethereal Sulfates
These are formed only by combination with phenolic hydroxyl groups, the product being an aryl sulfuric acid, R-OSO 3H. This reaction accounts for a considerable portion of the combined morphine excreted by dogs[1] . Epinephrine not oxidized in the tissues is excreted as a sulfuric ester, which may be epinephrine-(3 or 4)-hydrogen sulfate. Phenylethylamine and most other amines are oxidized by amine oxidases, but in the case of epinephrine conjugation is more rapid than oxidation and is therefore the dominant reaction [1] .
Glycine Conjugates
The formation of hippuric acid from benzoic acid in the organism is the best-known example of this reaction, which consists of the combination of glycine with an aromatic acid to form an aroyl glycine. The reaction takes place with carboxyl groups attached to the nuclei of benzene, naphthalene, thiophene, furane, and pyridine, the carboxyl group of the phenylacetic acids, and carboxyl groups separated from the aromatic nuclei by vinyl groups. In this class is the conversion of nicotinic acid and nicotinamide to nicotinuric acid, C 5H 4NCONHCH 2COOH, which, in the horse, is the main excretion product of nicotinic acid. This is of special interest because nicotinamide is a metabolite of nike-thamide [3] , [4] .
Acetylation
Acetylation in the body occurs only with aromatic amino groups and the amino groups of alpha amino acids. The amino groups of sulfonamides are largely acetylated, as are those of p-aminobenzoic acid and esters like procaine.
|
NH 2C 6H 4SO 2NH 2→ |
CH 3CONHC 6H 4SO 2NH 2 |
|
Sulfanilamide |
p-Acetaminosulfanilamide |
Methylation
Although probably a common reaction in the organism, methylation, from the viewpoint of detoxication mechanisms, is confined to the heterocyclic nitrogen of the pyridine and quinoline rings. In man nicotinamide is methylated to Nl-methylnicotinamide and excreted as such [5] . Large doses of nicotinamide also result in the excretion of nicotinuric acid, as in the horse [6] .
Drug Metabolism Variations among Different Animal Species
Considerable specific information is available about the fate of many drugs after administration to laboratory animals such as the dog, guinea-pig, rat and rabbit. Much is known about drug metabolism in man, of course, because most pharmacological research is designed to secure information that can be applied in human medicine
The absorption of drugs by the ruminant and the simple-stomached animal is not the same. Further, in the latter group carnivora and herbivora show differences, whereas omnivorous animals may show similarities to both. Absorption in the short digestive tract of the dog is relatively more efficient than in the longer digestive tract of the horse, which is adapted to forage and grains. In the case of ruminants certain drugs may be destroyed in the rumen or may be so mixed with indigestible fibre as to pass through the rumen digestive tract largely unabsorbed. The presence of enzymes in certain species and their absence in others account for variations between species. For instance, because of the presence of atropinase in the liver of the rabbit, atrophine is decomposed more readily than in other animals, notably swine [7] . Procaine is almost completely hydrolysed in man, but in the horse hydrolysis is much less complete [8] . In man, the rat, and the guinea-pig, N 1-methylnicotinamide is an important metabolite of nicotinic acid. It does not appear in the horse, rabbit, and sheep [1] , [9] .
Bearing in mind that such differences exist, examination of existing data does, nevertheless, show that animals belonging to the herbivorous group metabolize drugs similarly, and the same is true for carnivorous and omnivorous animals. Further, there are general similarities among all three classes which permit limited analogies to be drawn when data on certain species only are available. Information has been collected about the fate of drugs in all species that have been studied, and will be given in detail under each drug as the data are presented.
Relation of Chemical Structure to Metabolism
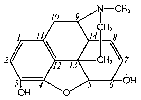
This important drug has been investigated by many workers, but definite information about the metabolic changes which take place after its ingestion has been forthcoming. only within relatively recent years. It is a phenanthrene derivative with two additional heterocyclic rings, one of which includes the tertiary amine nitrogen that gives the substance its basic properties. Its structure likewise includes one secondary, alcoholic hydroxyl group (position 6) and the phenolic hydroxyl group (position 3).
Oxidation at the nitrogen atom with removal of the methyl group has been demonstrated by the administration of morphine having carbon 14 in the methyl group attached to the nitrogen to both rats [10] and human subjects [11] . In both, C 14O 2 was detected in the expired air.
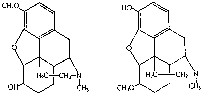
Both hydroxyl groups may undergo conjugation. Oberst [2] investigated the urinary excretion in morphine addicts of the following derivatives of morphine: 3,6-dia-cetylmorphine (heroin), alpha-isomorphine, 3-methylmorphine (codeine), dihydroheterocodeine, dihydrocodeine, dihydroisocodeine and dihydrocodeine methyl ether. The amounts of these substances, both free (unconjugated) and bound (conjugated), found in the urine after administration were determined and compared with similar results for morphine itself. Morphine, alpha-isomorphine, 3-methylmorphine, and dihydroheterocodeine were excreted in both free and bound forms. The bound fraction was two to ten times greater than the free in every instance. 3,6-diacetylmorphine (heroin) was excreted as morphine in both forms. Dihydrocodeine and dihydroisocodeine were excreted mainly in the free form, only small amounts being bound. Dihydrocodeine methyl ether was excreted only in the bound form. Thus, conjugation occurred only when either one or none of the hydroxyl groups was methylated. No conjugation occurred when both were methylated. Differences were noted between the stereoisomers, morphine and alpha-isomorphine and dihydrocodeine and dihydroisocodeine, giving rise to the speculation that molecular spatial arrangement affects the conjugation. No reason was apparent for the conjugating reaction being diminished in the hydrogenated codeine compounds. The 3,6-di-acetylmorphine was apparently first diacetylated and then metabolized as morphine. No evidence was found that a methyl group on either the phenolic or alcoholic hydroxyl group was altered or split off by the body. -
 | |
|---|---|
|
Dihydrococdeine |
Dihydroheterocodeine |
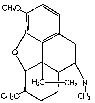
N-allylnormorphine, which differs from morphine only in the alkyl group attached to the nitrogen, is conjugated by surviving liver slices at the same rate as is morphine [12] .
Absorption of Morphine
Morphine is rapidly absorbed from all mucous surfaces. It is readily absorbed from the small intestine, and some may be absorbed from the stomach. It is absorbed promptly following subcutaneous injection. It is not absorbed through the intact skin, but a scarified epithelium permits slow entrance to the circulation [7] . It passes from the blood into the tissues, where it is temporarily stored [13] .
Dog
A dog begins to eliminate morphine in the urine and fèces immediately after absorption [7] . Much of the early work on the elimination of morphine after administration to dogs and other animals was subject to uncertainties inherent in the analytical methods employed [13] , [14] . The proportions of the drug excreted in the urine and the faeces were uncertain, and widely-varying results were reported by many workers. Nevertheless, the drug was found in both excretions consistently. Using improved methods, Wolff, Riegel, and Fry [14] found that at any level of single or repeated doses of morphine between 2 and 200 mg. normal dogs excreted, on the average, 20% of the drug. Two-thirds of the excreted morphine was recoverable in the urine and one-third in the fèces. None was found in saliva. Liaci[15] reported that 3.92% to 5.61% of morphine injected hypodermically into dogs was eliminated in the faeces. According to Inoue [16] a ten-milligramme quantity of morphine hydrochloride incubated with a ten-gramme portion of dog-liver slices can be recovered to the extent of 52% to 82%.
Gross and Thompson [17] , [18] showed that morphine was excreted in "free" and "combined" forms by both habituated and non-habituated dogs. Using suitable hydrolysis methods, total recoveries from non-tolerant dogs of 80% to 92% of the amount administered were obtained. From habituated dogs 35% to 66% was recovered. The" combined morphine" in the urine of dogs treated with morphine could be separated into easily and difficultly hydrolysable portions. Tolerant dogs excreted relatively greater amounts of the easily hydrolysable fraction, and non-tolerant dogs excreted much more of the difficultly hydrolysable fraction. Liver damage induced by carbon tetrachloride affected the conjugation of injected morphine [19] . The total morphine recoverable in the urine was not changed, but the easily hydrolysable fraction of conjugated morphine was decreased, and free morphine was proportionately increased.
The exact nature of the conjugated forms of morphine excreted by the dog is uncertain. According to Williams [1] the work of Stolnikow in 1884 indicated that in dogs the drug formed an ethereal sulfate. Gross and Thompson [17] reported that the easily hydrolysable form is hydrolysed at pH 1 in 60 minutes at a temperature of 100°C. and accounts for 30% of the combined morphine excreted by non-tolerated dogs. It is apparently not a glucuronide. It may be an ethereal sulphate in which the sulphuric acid group is attached to the phenolic 3-hydroxyl group [1] .
The following results seem to disagree with these suggestions. Seibert, Williams and Huggins [20] made a study of the urine from a dog which had been infused with 4 grammes of morphine sulphate. The free and bound morphine were separated chromatographically on paper with butyl alcohol-acetic acid-water after removal of urea with urease and concentration. Munier's alkaloid spray yielded two spots, one corresponding to morphine. The other was extracted with methanol and distilled water, the residue recrystallized, hydrolysed with betaglucuronidase and chromatographed on paper. The chromatogram indicated the presence of morphine, glucuronolactone, and glucuronic acid. An additional unknown spot was observed.
Morphine sulfuric ether given parenterally to tolerant and non-tolerant dogs is excreted in the urine as both free and bound morphine, but in lower amounts than reported for morphine itself [21] .
Woods [22] , [23] reports that the distribution of free and bound morphine in dogs and rats after administration of the drug is selective for certain tissues and fluids. Free morphine predominates in spleen, pancreas, thyroid, and adrenal tissues, but in lung, liver and kidney both free and bound forms are found. Very much higher concentrations are found in bile from the gall bladder than elsewhere. Brain, cerebrospinal fluid, and fat contain very little in either form. Biliary morphine is the principal source of faecal morphine.
Bound morphine from the bile is only a partial source of the morphine excreted in the urine in the first 24 hours after administration, but it is probably the source of all the bound morphine that is excreted in the urine after 24 hours. One abundant form of bound morphine in dog urine can be identified, after separation by chromatography, as the phenolic group-substituted monoglucuronide. Another form of bound morphine appears to be a different glucuronide of uncertain composition.
Man
Much of the work on the metabolism of morphine in humans has been done in connexion with studies of subjects addicted to the drug. In these an approximate average of 30% (11% to 55%) of varying doses of injected morphine is excreted in the urine, 5% free and 25% conjugated [24] , [2] . The remaining 70% is unaccounted for. It may be oxidized or eliminated as unidentified degradation products. That morphine is excreted as a conjugate with glucuronic acid is evident because the glucuronic acid output rises in proportion to the dose [2] . Conjugation can occur at either the 3- or 6-hydroxyl group [2] , but the exact nature of the glucuronide formed has not been determined. When the administration is oral, the amount excreted in the free state is less and the amount excreted combined is greater than when the administration is parenteral. Morphine is excreted in the urine of opium addicts [25] , [26] , and is found both "free" and "bound" in the urine of morphine addicts [24] , [27] . The faeces, liver, and perspiration of human addicts contain free morphine, bile contains only conjugated morphine, and urine and gastric contents contain both free and conjugated morphine. No morphine was found in saliva and blood [27] .
In a case of lethal poisoning from the subcutaneous administration of morphine the drug was found only in the urine (5 mg./100 ml. before hydrolysis and 17.7 mg./100 ml. after hydrolysis). None was found in the liver, blood, stomach contents, and intestines. None could be found in the liver even after strong hydrolysis [28] .
According to Autenreith [29] morphine is only slightly decomposed in cadavers even during putrefaction lasting for a long time. In the stomach and contents of an adult who took 25 grammes of tincture of opium and died four hours later, he found 0.028 gramme of nearly pure morphine. The drug was also found in the urine.
Morphine sulphuric ether given parenterally to addicts, non-addicts, and post-addicts is excreted as both free and bound morphine [21] .
More recent studies have been made with non-addicts of the excretion of carbon 14 (from the N-methyl group) after administration of single therapeutic doses of labelled morphine [11] . The major portion of the radioactivity is excreted in 24 hours. Urinary excretion accounts for most of the recovered activity, and faecal excretion accounts for 7% to 10% of the doses (intestinal excretion apparently being less important in man than in the rat). Pulmonary excretion of C[14] O 2 ranges from 3% to 6% of the injected dose in 24 hours.
Traces of morphine can usually be found in saliva and sweat after administration [29] , [30] , [31] . According to Munch [32] and Sollmann [13] Rosenthal appears to have been the first to demonstrate that morphine is eliminated in the saliva. It was consistently found in the saliva of hospital patients who were given therapeutic doses. Amounts ranged from 0.05 to 0.2 mg. of morphine.
Rat
Rats excrete the greater part of a dose of morphine in conjugated form [33] . In those which have developed tolerance the amount of conjugated morphine in the urine is decreased, but the free morphine remains the same. Liver slices from morphine-tolerant rats conjugate more morphine than liver slices from non-tolerant rats. The amount conjugated by liver sections from either is from 37% to 50% of morphine added [34] . No oxydimorphine is found in the tissues of either normal or habituated rats after administration of the drug [34] . The development of tolerance is not due to metabolism of the morphine in the liver, spleen, or gastro-intestinal tract [35] . Incubation of morphine with rat-liver slices followed by extraction of the material, boiling the solution in acid for thirty minutes, and analysis for morphine shows a sizeable loss. This was first attributed to oxidation by the rat liver [36] . However, autoclaving the solution at 20 pounds gauge pressure enabled quantitative recovery of the morphine and proved that, when incubated with rat liver, it is conjugated, not oxidized [37] . Also, the conjugation is effected in the absence of sulfate ion, making it probable that the conjugate is a glucuronide, not an ethereal sulfate.
After the administration of labelled morphine (carbon 14 in the N-methyl group) [10] to rats, the greatest concentration of radioactivity occurs in the urinary and intestinal tracts. Only minimal amounts appear in the central nervous system. Urinary excretion predominates, but there is evidence of an enterohepatic circulation of morphine or its metabolites following initial excretion into the bile. Male rats excrete significantly more C 14O 2 via the pulmonary route than females. Liver slices from male and female rats liberate C 14O 2 from labelled morphine, and the recoveries follow the same pattern as those for the in-vivoexperiments. It is suggested that some morphine is probably converted to normorphine, which may be excreted either free or conjugated. Similar demethylation has been observed with codeine [38] and with meperidine [39] . Tolerant animals do not demethylate more morphine than normal animals.
Rabbit
Normal rabbits after single subcutaneous injections of 50 to 200 mg./kg, of morphine hydrochloride eliminate more in the urine than in the faeces. Elimination is nearly complete within two days, and the total excretion ranges from 7% to 11%, the remainder being destroyed in the organism [40] . Liver slices conjugate morphine [37] . A ten-milligramme quantity of morphine hydrochloride incubated with a 10-gramme portion of rabbit liver slices is recovered to the extent of 48% to 51°/o [16] .
Horse
In experimental administrations of morphine to horses, both oral and parenteral, the drug has been detected in both saliva and urine [41, 42, 43, 44, 45] . It is frequently found in saliva after parenteral administration [42] . This indicates that actual excretion of the drug in the saliva does take place. The appearance of it in saliva after oral administration can conceivably be attributed to retention of some of the drug in the mouth, which is not possible, of course, when it is given parenterally.
Other Species
Normal cats after a single subcutaneous dose of 50 mg./kg. eliminate 11% to 27%, mostly in the Urine. Normal hens after a single subcutaneous dose of 100-200 mg./kg. eliminate 10% to 13% [40] .
Ten-milligramme quantities of morphine hydrochloride incubated with 10-gramme portions of liver slices from various animals at room temperature for thirty minutes can be recovered in the following percentages: cat, 50 to 74; guinea pig, 48 to 57; pigeon, 43 to 60; hen, 41 to 58 [16] .
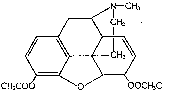
Relation of Structure to Metabolism
Heroin (3,6-diacetylmorphine) is readily hydrolysed to morphine. In alkaline solution (0.5 M Na 2CO 3at 26°C.) about 58% is hydrolysed in 2 minutes and 88% in 10 minutes [46] . Metabolism in the animal body involves, first, hydrolisis and, second, metabolism of the morphine produced by the hydrolysis.
Rabbit
Rabbit serum de-acetylates heroin. However, differences in the extent of the hydrolysis by individual animals have been demonstrated. In experiments using the sera from six male albino rabbits, Wright [47] observed that the sera of three of them liberated acetic acid (measured by the liberation of CO 2 from sodium bicarbonate) corresponding to 85% of the theoretical amount for hydrolysis of both acetyl groups. Sera from the other three animals hydrolysed the heroin more slowly and liberated acetic acid corresponding to 85% of the theoretical amount for only one acetyl group. Repeated observations confirmed these results. 6-monoacetylmorphine was then subjected to action by the rabbit sera, and it was found that the sera which could hydrolyse both acetyl groups of heroin were also able to hydrolyse the 6-acetyl group of the 6-monoacetylmorphine. Sera from the other three rabbits were not. Therefore, the serum from each of the rabbits was able to remove the 3-acetyl group of heroin, but that of only three could remove both groups. Sera from each of the six rabbits liberated acetic acid from diacetyldihy-dromorphine equivalent to 100% of the theoretical amount for one acetyl group. The sera from the three rabbits unable to de-acetylate 6-monoacetylmorphine hydrolysed diacetyl-dihydromorphine at a much slower rate. No serum was found which was able to remove the 6-acetyl group from the latter compound [48] .
The tissue found to have greatest enzyme activity was liver, followed in approximate order by kidney, brain, blood serum, and muscle. There was some evidence that the enzyme which catalyses the hydrolysis of atropine also catalyses the hydrolysis of heroin in the 6-position. There was also evidence that cholinesterase was not involved in the de-acetylation of heroin at either the 3- or 6-position [49] .
Ellis [50] found that the activities of rabbit plasma and liver were due to three distinct esterases. The monoacetylmorphine esterase, specific for the 6-acetyl group, was also found in the sera of man, dog, sheep, and guinea-pig. It was not present in the corpuscles. Most sera hydrolysed the 3-acetyl group of heroin, and the hydrolysis of this group appeared to be a function of the tributyrinase but not of the enzymes that hydrolyse methyl butyrate, acetylcholine, or acetylsalicylic acid.
Man
Human sera and tissues contain enzymes which de-acetylate heroin and dihydroheroin (diacetyldihydromorphine)[47] , [48] , [49] . The rate of de-acetylation for human serum is distinctly slower than for rabbit serum [47] , but in the body heroin seems to be completely hydrolysed [46] . In addicts physical dependence on morphine is satisfied by heroin, and the ratio of physical-dependence-satisfying doses of morphine sulfate and heroin hydrochloride is 2:1. About 7% of either drug is excreted in the urine as free morphine and about 50% as conjugated morphine.
Horse
Heroin is decomposed by horse serum. It is assumed that an esterase is the active agent and that one or two moles of acetic acid are formed [51] .
Heroin can be detected in saliva (as morphine) after administration to horses. This was proved in early experiments designed to establish the value of saliva as a medium for the detection of doping [52] . This author did not give details of the identifying tests, but it may be assumed that morphine, not heroin itself, was the substance actually present. The drug can be detected (as morphine) in both saliva and urine from horses after oral administration of 2.5 to 3 grains of heroin hydrochloride and after parenteral administration of 0.5 to 0.75 grains [42] . Samples of blood taken after two parenteral administrations proved to be negative.
Other Species
Sera and tissues from dogs, guinea-pigs, rats, and sheep have been shown to hydrolyse heroin [50] . The degree of activity varies from species to species[49] .
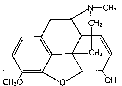
Relation of Structure to Metabolism
Codeine (3-methylmorphine) has only the 6-hydroxyl group available for conjugation. Such conjugation has been shown to occur in man [2] . Demethylation of the 3-methoxy group with the formation of morphine has been demonstrated [53] , [54] , [55] , as well as demethylation of the N-methyl group with the formation of norcodeine [38] .
Man
Oberst [2] found that the urinary excretion of codeine in morphine addicts after substitution [24] for morphine amounted to 11.2% of the dose in the free form and 31.6% in the conjugated form. The total amount excreted was slightly greater than that for morphine. Unaltered codeine conjugate was not extractable with an organic solvent. The free drug could be removed from the urine of an addict, the remaining urine subjected to acid hydrolysis and codeine detected in the hydrolysate.
Sollmann [13] states that codeine is excreted as such in the urine without demethylation. There is, however, more recent evidence that it is demethylated to morphine in man, free morphine being isolated from the urine of human subjects given 130 mg. of codeine orally in divided doses [55] . After an oral administration of codeine phosphate in man, there was found a considerable quantity of an unknown metabolite, in addition to previously observed metabolites - that is, morphine and conjugated codeine base [54] . When subjected to counter-current distribution studies, this substance proved to have characteristics identical with those of N-demethylated codeine (norcodeine). Only part of the norcodeine excreted appeared in the free form, hydrolisis under 20 pounds pressure being required to recover the remainder [38] .
Dog
Dogs are reported to excrete about 80% of administered codeine, mainly in the urine but some in the fèces. Repeated administration does not increase its destruction (13).
Sanfilippo [56] , [57] , [58] found that injections of 0.15 to 0.5 mg./kg. of morphine produced hyperglycemia in dogs. Daily injections of 5 mg./kg. of codeine for one week caused habituation, so that 0.5 mg./kg. of morphine no longer caused hyperglycaemia. This was considered evidence that codeine is demethylated in vivo by the dog.
Rat
Demethylation by the rat, in vivo and in vitro, of codeine labelled with C 14H 3 at the 3-position has been demonstrated [53] . The methyl group is oxidized and eliminated as C 14O 2. The in vitro studies showed that the liver was the most important site for the demethylation. Comparison of residual codeine results by the methyl orange technique [59] with results obtained by measuring carbon 14 radioactivity showed good agreement. Thus the production of C 14O 2 was probably the result of demethylation rather than transmethylation. Demethoxylation was not definitely ruled out, but Bernheim [36] showed that 13% of codeine incubated with liver slices appeared as a phenolic-OH compound, and it is probable that the process is one of demethylation. After a subcutaneous injection of 40 mg./kg. of the labelled codeine, approximately one-fifth of the radioactivity appeared as C 14O 2 in the expired air within 2 hours, 40% within 6 hours. Latham and Elliot [60] reported C 14O 2 in the expired air of rats administered carbon 14-methoxy-labelled codeine. Codeine was excreted unchanged by the kidney and also into the lumen of the gastro-intestinal tract. The liver was found to metabolize the labelled codeine to a substance with solubility characteristics different from those of codeine itself. This unidentified radioactive substance was excreted into the duodenum via the biliary tract. The liberation of morphine from codeine by incubation with rat-liver slices was demonstrated by Adler and Shaw [54] . The total amount of morphine formed represented less than half of the metabolized codeine, and the liberated morphine was further metabolized by conjugation. Identification of the morphine was effected by means of the ultraviolet spectrum of the recovered metabolite, preparation of the 2,4-dinitrophenyl derivative, and comparison of its crystal form, ultraviolet spectrum, and X-ray diffraction pattern with those of the known morphine derivative. After subcutaneous injection of codeine labelled with carbon 14 in the N-methyl group (4 mg./kg.) about 13% of the amount given appeared as C 14O 2in the expired air [45] .
Horse
After an oral administration of 30 grains of codeine phosphate the drug was detected in both the urine and saliva[45] .
WILLIAMS, R. T.: Detoxication Mechanisms, Wiley (1947).
002OBERST, F. W. : J. Pharmacol. Exptl. Therap., 69, 240 (1940).
003ELLINGER, P.; ABDEL KADER, M. M.: Biochem. J., 44, 77 (1949).
004COULSON, R. A.; STEWARD, C. A.: Proc. Soc. Exptl. Biol. Med., 61, 364 (1946).
005HUFF, J. W.; PERLZWEIG, W. A.: J. Biol. Chem., 150, 394 (1943).
006JOHNSON, B. C.; HAMILTON, T. S.; MITCHELL, H. H.: ibid., 159, 231 (1945).
007JONES, L. M.: Veterinary Pharmacology and Therapeutics, Iowa State College Press (1954).
008BRODIE, B.B. : Bulletin No. 80, Association of Official Racing Chemists (1949).
009JOHNSON, B. C.; LIN, Pei-Hsing : J. Am. Chem. Soc., 75, 2971 (1953).
010MARCH, C. H.; ELLIOT, H. W.: Proc. Soc. Exptl. Biol. Med., 86, 494 (1954).
011ELLIOT, H. W.; TOLBERT, B. M.; ADLER, T. K.; ANDERSON, H. H.: ibid., 85, 77 (1954).
012SEIBERT, R. A.; HUGGINS, R. A.: ibid., 82, 518 (1953).
013SOLLMANN, T.: Manual of Pharmacology, 6th Ed., Saunders.
014WOLFF, W. A.; RIEGEL, C.; FRYSE, E. : J. Pharmacol. ExptI. Therap., 4 ; 391 (1933).
015LIACI, L.: Arch. Farmacol. Sper., 72, 38 (1941).
016INOUE, T. : Japan J. Med. Sci. IV, Pharmacol., 12, No. 283; Proc. Japan. Pharmacol. Soc., 13, 75 (1940).
017GROSS, E. G.; THOMPSON, V. : J. Pharmacol. Exptl. Therap., 68, 413 (1940).
018GROSS, E. G.; THOMPSON, V.: ibid., 72, 138 (1941).
019GROSS, E. G.: Proc. Soc. Exptl. Biol. Med., 51, 61 (1942).
020SEIBERT, R. A.; WILLIAMS, C. E.; HUGGINS, R. A.: Science, 120, 222 (1954).
021OBERST, F. W. : J. Pharmacol. Exptl. Therap., 80, 188 (1944).
022COCHIN, J.; HAGGART, J.; WOODS, L. A.; SEEVERS, M. H.: ibid., 111, 74 (1954).
023WOODS, L. A.: ibid., 112, 158 (1954).
024OBERST, F. W.: ibid., 69, 240 (1940).
025CHOPRA, R. N.; CHOPRA, G. S.; ROY, A. C.: Indian J. Med. Research, 29, 195 (1941).
026LIANG, C. K.: Natl. Med. J. China, 22, 507 (1936).
027OBERST, F. W. : J. Pharmacol. Exptl. Therap., 74, 37 (1942).
028ZWIKKER,J. J. L.; STEENHAUER, A.J.: Pharm. Weekblad , 79, 241 (1952).
029AUTENREITH, W.: Laboratory Manual for Detection of Poisons and Powerful Drugs, Tr. Warren, W. H., 6th Ed., Blakiston (1928).
030KRUEGER, H.; EDDY, N. B.; SUMWALT, M.: U.S. Public Health..Service, Suppl. No. 165, 1 (1941); 813 (1943).
031MILKS, H. J.: Veterinary Pharmacology, Materia Medica and Thera-peutics, 6th Ed., Eger (1949).
032MUNCH, J . C. : J. Am. Pharm. Assoc., 23, 1185 (1934).
033ZAUDER, H. L. : J. Pharmacol. Exptl. Therap., 104 , 11 (1952).
034FICHTENBERG, D. G. : Bull. Narcotics , U.N. Dept. Social Affairs, 3, No. 4, 28 (1951).
035FAWAZ, G. : Proc. Soc. Exptl. Biol. Med., 68 , 262 (1948).
036BERNHEIM, F.; BERNHEIM, M. L. C. : J. Pharmacol. Exptl. Therap., 81 , 374 (1944).
037BERNHEIM, F.; BERNHEIM, M. L. C. : ibid., 83 , 85 (1945).
038ADLER, T. K. : ibid., 106 , 371 (1952).
039PLOTNIKOFF, N. D.; ELLIOT, H. W.; WAY, E. L. : ibid., 104 , 377 (1952).
040YOSIKAWA, N. : Japan. J. Med. Sci. IV, Pharmacol ., 12, No. 283; Proc. Japan. Pharmacol. Soc., 13 , 74 (1940).
041JAGGARD, G. : Proc. 3rd AORC Meet ., App. XXIII, page 8 (1949).
042MORGAN, C. E.: Proc., 1st AORC Meet ., 16 (1947).
043MORGAN, C. E. : Proc., 5th AORC Meet ., App. XX, 3 (1951).
044OESTER, Y. T. : Proc., 1st AORC Meet ., 14 (1947).
045OWENS, F. P. : Proc., 3rd AORC Meet ., App. XXVII, Table 2 (1949).
046OBERST, F. W. : J. Pharmacol. Exptl. Therap ., 79, 266 (1943).
047WRIGHT, C. I. : Science, 92 , 244 (1940).
048WRIGHT, C. I. : J. Pharmacol. Exptl. Therap., 71 , 164 (1941).
049WRIGHT, C. I.: ibid., 75 , 328 (1942).
050ELLIS, S. : ibid., 94 , 130 (1948).
051MASSART, L.; DUFAIT, R. : Naturwissenschaften, 29 , 572 (1941); Chem. Zentr., 1 , 496 (1942).
052FAILLE, J. : Recherches chimiques, physiologiques et toxicologiques sur labrucine , Thesis, National Veterinary School, Alfort; Faculty of Medicine, Paris (1928).
053ADLER, T. K.; LATHAM, M. E. : Proc. Soc. Exptl. Biol. Med., 73 , 401 (1950).
054ADLER ,T. K.; SHAW, F. H. : J. Pharmacol. Exptl. Therap., 104 , 1 (1952).
055MANNERING, G. J.; DIXON, A. C.; BAKER, E. M.; ASSAMI, T.: ibid., 111 , 147 (1954).
056SANFILIPPO, G. : Bull. Soc. Ital. Biol. Sper., 23 , 1119 (1947).
057SANFILIPPO, G. : ibid., 24 , 723 (1948).
058SANFILIPPO, G. : ibid., 29 , 1761 (1953).
059BRODIE, B. B.; UDENFRIEND, S. : J. Biol. Chem., 158 , 705 (1945).
060LATHAM, M. E.; ELLIOT, H. W.: J. Pharmacol. Exptl. Therap., 101 , 259 (1951).


