Introduction
Materials and methods
Results
Discussion
Acknowledgements
Author: R. E. GAREY, R. G. HEATH, C. E. TURNER
Pages: 55 to 66
Creation Date: 1980/01/01
Several published reports have suggested that prolonged or heavy use of the crude drug marihuana may result in observable psychological and behavioural changes (1, 2, 3). A theoretical basis for these changes could be effects on brain structures and functions caused by constituents in marihuana smoke. To determine if such effects do occur when marihuana is smoked or when pure (-)Δ [ 9] - trans-tetrahydrocannabinol (Δ [ 9] -THC) is administered, a series of long-range experiments were designed to investigate the effects of acute and chronic marihuana smoke on electroencephalograms and structure integrity of certain brain areas. Previously we have reported our findings on electroencephalograms (4, 5, 6) and the integrity of brain areas (7, 8).
In this communication, we wish to report and discuss procedures used in our programmes; to provide detailed information on methodologies; and to compare the blood levels of cannabinoids found in monkeys after smoking marihuana; to compare blood chemistry of monkeys after smoking marihuana or a placebo, and in general present data that hopefully will stimulate studies with marihuana using the rhesus monkey model for obtaining biological data.
The marihuana to be smoked was weighed and packed into a brier pipe and attached to a modified respirometer (5). The marihuana was then lit and the smoke drawn into the respirator by a variable-speed electric motor until combustion of the marihuana was completed. The smoke was delivered through a tube connecting the machine to the nasopharynx of the monkey at a rate and time commensurate with the rate and time observed in humans.
If the smoke from the machine was to be used for analysis rather than delivery to a monkey, identical conditions were employed, except that the plastic tube from the machine was connected to a metal tube inserted into a 25 ml glass vial that contained 15 ml of 95 per cent ethanol. The metal tube, to which the plastic tube from the machine was attached, was inserted into the vial without touching the bottom of the vial. An additional shorter metal tube which was inserted through the rubber stopper, but not immersed in the ethanol, served as a vent to allow the pressure, built up by the smoke and air delivered from the machine, to escape once the smoke had passed through the ethanol. The ashes and non-combusted residue were removed from the pipe and stored for subsequent analysis.
Additional experiments were performed in which the plastic tubing from the machine was replaced by a glass tube which was inserted through the top of the rubber stopper directly into the ethanol. In some cases the glass tube was passed from the machine directly into ethanol and tile vial sealed with a glass stopper immediately after delivery of the smoke.
Human subjects smoked similar amounts of marihuana to that burned by the machine. The subjects, after taking a deep inhalation on tile pipe and without passing the smoke into their lungs, would expel the smoke into a plastic tube of the same length and composition as that used to connect the monkeys to the machine. The tube was connected to a glass-stoppered vial containing 15 ml of 95 per cent ethanol as described above. The ashes were removed from the pipe and stored for analysis.
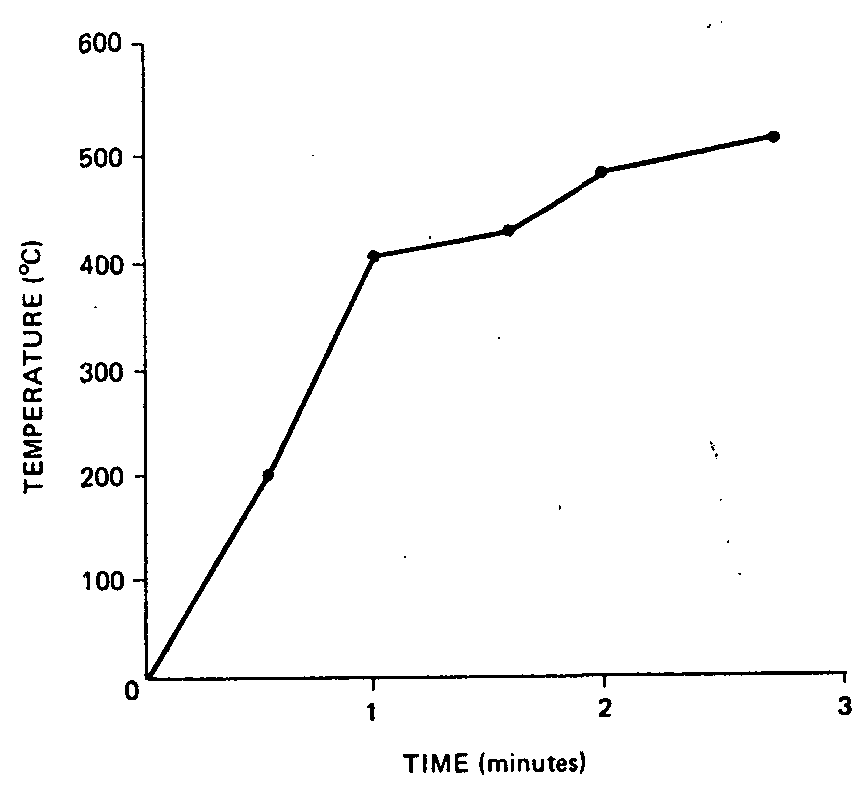
A Cole-Parmer model 8396 electric pyrometer, equipped with a model 8396-22 thermocouple, was used in these experiments. The thermocouple was inserted before combustion into the pipe and secured so that continuous temperature recordings could be made throughout the combustion of a predetermined amount of marihuana (fig. 1).
The vial containing Cannabis smoke trapped in 15 ml of 95 per cent ethanol was ultrasonically shaken for 30 seconds, after which time, the solution was decanted into a round-bottomed flask of known weight. The vial was washed with an additional 15 ml of 95 per cent ethanol which was also decanted into the round-bottomed flask. Ethanol was then removed in vacuo at ambient temperature. Residue weight was then determined by reweighing the flask.
At this point, 0.10 ml of absolute ethanol containing 1.0 mg of androst-4-ene-3,17-dione was added as the internal standard. Routinely, 0.3 μl of the resulting solution was injected with 0.2 μl of ethanol as a flush solvent.
Analyses were performed with gas chromatographs (Beckman GC-72-5 and GC-65) equipped with hydrogen flame-ionization detectors and operated isothermally at 210°C. Inlet and detector temperatures were 240° and 260°, respectively. Glass columns, 0.64 cm (0.25 in.) o.d. and 2 mm i.d. x 2.43 (8 ft) were packed with 2 per cent OV-17 or 6 per cent V-1 on 100-200 mesh Chromosorb WHP or 100-120 mesh Gas Chrom Q. Nitrogen was used as the carrier gas at a flow rate between 10 and 30 ml/min, depending upon instrument and column requirements. Usually the head pressure was between 26 and 40 psi.
Peak area measurements in millivolts were made with a computer (Digital, PDP-8). The concentration of each cannabinoid was obtained by dividing the total millivolts for all cannabinoid peaks into the millivolts for each peak. Milligrams present were then obtained by multiplying the total weight of the residue by the per cent of cannabinoids determined to be present in the sample. Since the sample size was so small, this procedure provided more consistent data than the routine method of comparing the area of each peak to the area of the internal standard and using response factors. Thus, the internal standard was used only to establish relative retention times for each peak.
Ashes and non-combusted residue were weighed and extracted with 40 ml of chloroform at ambient temperature for 1 hour. Solids were removed by filtration, and the mother liquor was concentrated in vacuo at ambient temperature. Ethanol containing the internal standard was then added. The procedure was basically the same as described for analysis of the ethanol solution. In this case, however, a dry weight and not a normalization analysis was obtained. Thus, the area of each cannabinoid peak, measured in millivolts, was compared to the peak area of the internal standard. The amount of internal standard added varied according to the weight of the sample.
The analytical procedures used are basically those reported by Turner and co-workers (9, 10).
A. Radioimmunoassays were performed by the Cannabinoid Quantification Serum Laboratory, White Memorial Medical Center, Los Angeles, California 90033;
B. Gas chromatography-mass spectrometry analyses were performed by Battelle Laboratories, Columbus, Ohio 43201.
For both analyses, several baseline blood samples were drawn from the monkeys before they began smoking and the plasma separated and stored at - 20 oC in glass vials with Teflon-lined caps. Naive animals, not previously exposed to marihuana, received smoke from various amounts of marihuana for one to four days. Plasma samples, obtained from the animal at 10 minutes post-smoking, were frozen and coded for subsequent analysis. Analysis by both laboratories were performed "blind" with the code being broken by Dr. Robert Willette at the National Institute on Drug Abuse. All animals were exposed to smoke from a marihuana cigarette containing 2.45 per cent Δ [ 9] -THC, 0.42 per cent CBN, 0.01 per cent Δ [ 8] -THC, 0.12 per cent CBD, and 0.20 per cent CBC.
Blood samples were drawn for laboratory analysis before exposure to marihuana and at various times after smoking. Urine was obtained at similar times. Analyses included:
Na+ and K+ levels in serum and urine
Creatine in urine
Creatinine in urine
Serum SGOT, CPK, and aldolose levels with use of materials and according to the Cal-Biochem Stat-Pacs.
A separate group of monkeys were injected intravenously with various amounts of pure (95 per cent) Δ [ 9] -THC obtained from NIDA. For injection, the Δ [ 9] -THC was prepared in the following manner: A 1.5 ml aliquot of lipemic human serum was mixed with the desired amount of a 100 mg/ml of 95 per cent ethanol solution of Δ [ 9] -THC. The mixture was sonicated cold with a Sonifier Cell Disrupter, Model W1859, equipped with a microtip. Output dial was set to position 3-4 for 5-10 seconds, or until a smooth emulsion was achieved. The test animals were then injected with the total aliquot.
The monkeys involved in these experiments were devided into different groups, designated by the type, frequency and amount of marihuana they received (see table 1).
|
Group |
Exposure |
Durations (months) |
mgΔ'-THC/joint |
|---|---|---|---|
|
1. Heavy active
|
3 x day, 5 days/week
|
6 | 23.80 |
|
2. Moderate active
|
1 x day, 2 days/week (T&T)**
|
6 | 23.80 |
|
3. Light active
|
1 x day, 5 days/week
|
6 | 6.50 |
|
4. Heavy inactive
|
3 x day, 5 days/week
|
6 | 0.02 |
|
5. Δ9-THC (pure)
|
1 x day, 5 days/week
|
6 | 0.70 |
*All experiments were performed using marihuana containing 2.45 per cent Δ 9-THC.
**T & T stands for Tuesday and Thursday.
Analysis of the smoke from the machines and from human subjects showed that the smoke from the humans contained about eight times more Δ [ 9] -THC than did the smoke from the machine. Results are compared in figure 2.
The decrease in Δ [ 9] -THC delivered in the smoke from the machine probably occurred due to absorption of the cannabinoid by the machine and tubes connecting the machine to the monkey. After each experiment, the machine and connective tubing were thoroughly cleaned with ethanol and acetone and were rinsed with water to avoid contamination of the next batch smoked. The material obtained from these washings was not analysed. Since the human smoke was obtained in the same manner, except for the passage of the smoke through the machine, it seems evident that passing and storing the smoke in the machine were the major sources of loss of cannabinoids in the smoke. Turner et al. (11) observed that 20 per cent of the cannabinoids are absorbed when metal columns were used in analytical procedures.
|
Human versus machine
|
||
|
Methods
|
Smoke (mg Δ9 - THC)
|
Ashes (mg Δ9 - THC)
|
|
Human (N=4)
|
6.4 per cent (25.10 per cent)
|
3.8(14.70 per cent)
|
|
Machine (N=12)
|
0.86 (3.42 per cent)
|
Trace (0.10 per cent)
|
|
Per cent recovery from 1 gram of marihuana containing 24.5 mg Δ9 - THC
|
||
|
Agurell el al., (22)
|
||
|
Procedure
|
Per cent Δ9 - THC in smoke
|
|
|
Humans smoking hashish
|
14-20 per cent
|
|
|
Humans smoking cannabinoids impregnated on tobacco
|
14-29 per cent
|
One batch of marihuana, obtained from local police authorities, which human smokers had reported to be very potent was investigated. Analysis of this marihuana (K-23) by methods previously described and comparison with another batch (44) raise some interesting points. Batch K-23 was relatively clean of minor cannabinoids. Batch 44, based solely on the Δ [ 9] -THC content, should have been about 2.58 times as potent as K-23. This was not found to be so experimentally, since qualitatively K-23 was very much more effective in eliciting behavioural and EEG changes in two experimental monkeys. This observation seems to substantiate the subjective human reports of potency of K-23 (see table 2).
|
CBDV |
CBD |
CBC |
CBGM |
Δ8-THC |
Δ-THC 9 |
CBN |
|
|---|---|---|---|---|---|---|---|
|
Batch 44
|
0.01 | 0.12 | 0.20 | 0.02 | 0.01 | 2.45 | 0.42 |
|
Batch K-23
|
-
|
-
|
0.10 |
-
|
0.04 | 0.95 | 0.63 |
CBDV-Cannabidivarin; CBD-Cannabidiol; CBC-Cannabichromene; CBGM-Cannabigerol Monomethylether; CBN-Cannabinol.
Analytically, K-23 contained only Δ [ 9] -THC, CBN, CBC and a very small amount of Δ [ 8] -THC. Breaking these percentages into mg/g (Δ [ 9] -THC=9.5, CBN= 6.3, CBC = 1.0 and Δ [ 8] -THC = 0.4) provides no absolute data as to why K-23 was more potent than the analysis indicated. However, Cannabis preparations that do not contain CBD, but that do contain CBC are regarded as the most potent (12). Karniol and Carlini and others (13, 14, 15, 16) have shown that, in laboratory animals and man, CBD interferes with the effects of Δ [ 9] -THC, and CBN potentiates the effects of Δ [ 8] -THC. In research of antagonism and potentiation of cannabinoids by other cannabinoids, no work has been done on the role of CBC. This is understandable since for many years CBC was misidentified as CBD(17). With the recent development of an excellent synthesis for CBC (18), studies can now be designed which may clarify the biological effects of CBC in marihuana.
Recent findings by Elsohly et al. (17) found mono- and dihydroxylated cannabinoids, with the double bond in the 6α-10α position, present in certain stored Cannabis samples (marihuana). These new cannabinoids are transformed into CBN when subjected to GLC analysis. In fact, many different hydroxylated cannabinoids have been found in the decomposition pathway of Δ [ 9] -THC to CBN (20, 21). Pharmacologic action of these new cannabinoids have not been determined, but from sterochemical aspects some should have Δ [ 9] -THC like activity. Since batches K-23 and 44 were stored under different conditions it is possible that certain new hydroxylated cannabinoids were present or available in different amounts in these batches; thus, possibly giving K-23 a more pronounced Δ [ 9] -THC like activity than would be expected from the analytical data.
The cannabinol peak reported for batch 44 was contaminated with a C 2 9 hydrocarbon whereas K-23 contained only CBN.
In addition to the group of monkeys given marihuana, another group was exposed to smoke at the same dosage level as the heavy active group using placebo material. Analysis of the placebo, marihuana from which the cannabinoids had been extracted, showed little, if any, Δ [ 9] -THC or Δ [ 8] -THC remained in the sample itself, or smoke obtained from combustion (table 3).
|
Material |
Δ8 |
Δ9 |
|---|---|---|
|
Placebo
|
0.02 | 0 |
|
Placebo smoke
|
Trace
|
0 |
|
Placebo ash
|
Trace
|
0 |
All the smoke samples analysed, including the placebo, showed quantities of a substance with a relative retention time of 0.44 using gas chromatography. This fact suggested that one of the cannabinols was being transformed into Δ [ 8] -THC, which has a relative retention time of 0.44, or that a contaminant was being introduced which chromatographed as Δ [ 8] -THC. It was unlikely that Δ [ 8] -THC was being formed since the placebo smoke also contained the same peak. The peak reported to be Δ [ 8] -THC was, therefore, investigated by gas chromatograph-mass spectroscopy (GC-MS) and proved to be composed of phthalates. The major phthalate was identified by GC-MS as dibutylphthalate. A sample of pure dibutylphthalate did, indeed, have a relative retention time of 0.44 and the GC-MS data were identical to the unknown.
To identify the source of phthalate contamination, experiments were performed in which glass vials and glass tubing rather than plastic were used to connect the machine to the ethanol trap. Results of these experiments indicate that neither the machine nor the tubing was the source of contamination. Only three possibilities remained as a source of phthalate: the pipe, the rubber stoppers used to hold the glass tube in the ethanol trap, or the rubber stopper used to cap the vials. Since the pipe was brier, it was eliminated. Rubber stoppers are known to contain phthalate, and dibutylphthalate was easily extracted by the ethanol used to trap the cannabinoids. However, since no rubber stoppers were utilized in the system employed to deliver marihuana smoke to the experimental monkeys, no phthalates were available to the monkeys and thus could not have affected the experimental results.
The temperature of combustion at which the machine burned marihuana was determined to be 400°C after one minute and 500°C after two minutes (fig. 1). Packing the pipe in ice to reduce the temperature nor slowing the total burning time affected Δ [ 9] -THC content in the smoke. Thus the extremely low amounts of Δ [ 9] -THC present in smoke from the machine was due to the configuration and materials of the machine. Per cent of Δ [ 9] -THC found in the smoke of human volunteers was within the range reported by Agurell et al. (22) for hashish (fig. 2).
Time required by human subjects to smoke a given amount of marihuana was between 15 and 20 minutes, thus the respirometer was calibrated to deliver smoke to the monkey in 17-20 minutes. This required storage of the smoke in the machine reservoir for that period. Thus, the smoke could be cooled and administered to the monkey at a constant temperature during the same time span as for humans. However, as previously discussed, this storage arrangement was found to be the source of the loss of Δ [ 9] -THC and other cannabinoids in machine smoke.
Plasma levels of Δ [ 9] -THC in monkeys obtained by radioimmunoassay and mass spectrometry analysis initially were not well correlated. The relative values do, however, show a pattern of increased Δ [ 9] -THC levels after marihuana smoking that would be expected based on the amount of marihuana smoked (table 4). For comparison, values obtained from naive human subjects and chronic marihuana users see references 23, 24, 25.
Table 5 is a summary of the biochemical studies on the blood and urine of monkeys exposed to marihuana smoke of Δ [ 9] -THC for one, three and six months. All values obtained were compared to: (1) pre-smoking values; (2) values obtained from placebo smoked animals; and (3) values obtained at 2 and 24 hours post-smoking. Data were analysed for significant differences by Students' T Test. Changes were not significant in any of the measurements except the 2 hour post-smoking creatinine and urinary creatine levels and the 24-hour post-smoking creatinine levels.
Table 4
|
Subject |
No. joints |
mg Δ 9-THC/joint |
Time post smoking (min) |
Plasma level ng Δ 9-THC/ml |
|---|---|---|---|---|
|
RIA
|
||||
|
Monkey (naive)
|
1 | 5.4 | 5 | 8 |
|
Monkey (naive)
|
1 | 21.0 | 5 | 35 |
|
Monkey (naive)
|
3 | 23.5 | 5 | 40 |
|
Human (naive)
|
1 | 10.0 | 10 |
25-50
|
|
Human (chronic)
|
1 | 10.0 | 10 |
200-250
|
|
Monkey (chronic)
|
1 | 5.4 | 5 | 30 |
|
Monkey (chronic)
|
1 | 21.0 | 5 | 53 |
|
Monkey (chronic)
|
3 | 23.5 | 5 | 76 |
|
GC-MS
|
||||
|
Monkey (naive)
|
1 | 21.0 | 5 | 3 |
|
Human (naive)
|
1 | 10.0 | 10 |
19-26
|
|
1.
|
No significant difference found between pre-smoking values and 1, 3 and 6 month post-smoking or post injection values for serum Na+, K+, CPK, SGOT, aldolase or urinary Na+, K+, creatinine, creatine.
|
|
2.
|
No significant differences found between values for placebo smoked animals and marihuana smoked of Δ
[
9] -THC injected animals at 1, 3 or 6 months for serum Na+, K+, CPK, SGOT, aldolase or urinary Na+, K+, creatine, creatinine.
|
|
3.
|
No significant difference found between placebo smoked and marihuana smoked Δ
[
9] -THC injected animals at either 2 or 24 hours post treatment for serum Na+, K+, CPK, SGOT, aldolase or urinary Na+, K+.
|
|
4.
|
Significant and non-significant findings of a comparison of the 2 and 24 hour urinary creatinine and creatine levels between placebo smoked and active smoked of Δ
[
9] -THC injected animals by groups.
|
|
A.
|
Increased creatinine 24-hour levels
|
|
|
Heavy active
|
P < 0.005
|
|
|
Moderate active
|
P < 0.025
|
|
|
Δ
[
9] -THC
|
P < 0.010
|
|
|
Light active
|
NS
|
|
|
B.
|
Increased creatinine 2-hour levels
|
|
|
Heavy active
|
P < 0.01
|
|
|
Δ
[
9] -THC
|
P < 0.01
|
|
|
Moderate active
|
NS
|
|
|
Light active
|
NS
|
|
|
C.
|
Increased creatine 2-hour levels
|
|
|
Heavy active
|
P < 0.005
|
|
|
Moderate active
|
P < 0.05
|
|
|
Δ
[
9] -THC
|
P < 0.005
|
|
|
Light active
|
NS
|
|
|
D.
|
Creatine 24-hour levels
|
|
|
All groups
|
NS
|
Although a significant decrease was observed in the Δ [ 9] -THC content of the smoke from the machine in comparison to the smoke generated by a human, the decrease is not sufficient to explain the relatively low Δ [ 9] -THC levels found in the plasma of monkeys as compared to those found in humans. Other conditions are apparently affecting plasma levels of Δ [ 9] -THC and preventing levels from reaching expected values. We attribute this effect to at least three factors: (1) inefficient delivery of the smoke to the animals, (2) failure of the monkeys to deeply inhale the smoke administered, and (3) expulsion of the smoke as rapidly as possible by the monkeys. Human subjects tend to take deep puffs on the pipe and to hold the smoke in their lungs, for a prolonged period allowing more time for absorption of the cannabinoids in the smoke. The monkeys, although continuously exposed to the marihuana smoke for 20 minutes, did not inhale the smoke as deeply as humans, and they tried to expel the smoke as quickly as possible. These two conditions would lead to decreased absorption of psychoactive components. The monkeys were exposed to the smoke for a greater period of time than humans, but plasma levels of Δ [ 9] -THC in the monkeys were lower than expected; thus, both the loss of Δ [ 9] -THC in the machine and the inefficiency of delivering the smoke appeared to be preventing a predictable rise in Δ [ 9] -THC levels in plasma based on a weight-to-weight (mg/kg) basis. Levels of Δ [ 9] -THC reached by chronic human smokers were far in excess of those obtained by our monkeys, even when they were exposed to smoke three times a day with the equivalent of one human "joint" each time. As previously mentioned, these levels suggest that higher volumes of marihuana smoke or marihuana with higher concentration of Δ [ 9] -THC and other cannabinoids must be smoked if the Δ [ 9] -THC levels in plasma of monkeys are going to reach Δ [ 9] -THC level comparable to those reached by human smokers. Alternatively, more effective methods of delivering the smoke to the monkeys must be developed.
Plasma Δ [ 9] -THC levels using the RIA procedure and using the GC-MS procedure should give different values. Radioimmunoassays are sensitive, but give cross-reactivity with many cannabinoids and cross-reactions with different antiserum also adds variability (26, 27, 28). The GC-MS procedure for Δ [ 9] -THC does not have the problem of "cross-reactivity" since only Δ [ 9] -THC's mass fragments are monitored (29). This can account for the differences in Δ [ 9] -THC in human naive subjects (table 4) receiving one joint of 10 mg. Gas chromatography - mass spectra data confirmed 19-26 mg/ml after 10 minutes post smoking time whereas with RIA the range was 25-50 mg/ml. Moreover, the very high 200-250 mg/ml in the chronic smoker obviously reflects the total "cannabinoid" content in blood and not just Δ [ 9] -THC.
Hollister (3) reported a reduction in the creatinine clearance of humans after smoking marihuana, but was not sure this temporary decrease was of any clinical significance. The increased urinary creatinine levels at 2 and 24 hours post-smoking and the increased creatine levels at 2 hours post-smoking are apparently due to the presence of Δ [ 9] -THC since both the active smoke groups and the Δ [ 9] -THC injected group showed the increase whereas the placebo smoked group did not. The significance of these findings is unknown. Data are given in table 5.
The amount of smoke delivered to monkeys in these experiments was similar to the amount delivered to a human smoking an equivalent amount of marihuana. To rule out the effects of excessive smoke, a control group of animals was exposed to smoke on the same schedule as the heavy marihuana smokers. In this experiment placebo marihuana was used. Since neither acute nor chronic effects of placebo were seen on the biochemical, electroencephalographic, histologic or behavioural phenomena being measured, the smoke itself did not affect the measurements, and changes reported seemed to depend solely on the cannabinoids.
The levels of Δ [ 9] -THC in the smoke and plasma, when marihuana was smoked, helped reduce a potential inherent problem: that the doses of marihuana smoke administered to the monkeys were reasonable and not excessive.
Moreover, failure of placebo to induce EEG, biochemical or neuro-anatomical changes, similar to those observed with marihuana and Δ [ 9] -THC, support the observation that the cannabinoids and not the smoke itself were responsible for the biological changes. Thus, the method developed to deliver marihuana smoke to monkeys was effective. Work is in progress on a more efficient delivery system which will give a more favourable ratio of cannabinoids per unit of smoke.
The authors are grateful to Ms. Peggy Carlisle for typing, and to Mrs. Glenda Lewis for running the chromatographic analyses. This investigation was supported by Grant Number R01 DA 00311, Contract Numbers HSM-42-70-109 and 271-78-3527 from the National Institute on Drug Abuse, The Research Institute of Pharmaceutical Sciences, School of Pharmacy, University of Mississippi and the Department of Psychiatry and Neurology, Tulane University Medical School, New Orleans, Louisiana.
F. S. Tennant and C. J. Groesbeck, "Psychiatric effects of hashish", Archives of General Psychiatry , vol. 27, 1972, pp. 133-136.
002H. Kolansky and W. T. Moore, "Effects of marihuana on adolescents and young adults", Journal of the American Medical Association , vol. 216, 1971, pp. 486-492.
003L. E. Hollister, "Marihuana in man: Three years later", Science, vol. 172, 1971, pp. 21-28.
004R. G. Heath, "Marihuana", Archives of General Psychiatry , vol. 26, 1972, pp. 577-584.
005R.G. Heath, " Cannabis sativa derivates: Effects on brain function of monkeys", in Marihuana: Chemistry, Biochemistry and Cellular Effects , G. G. Nahas, ed. (New York, Springer-Verlag, 1976), pp. 507-519.
006R. G. Heath, "Marihuana and Δ 9-THC: Acute and chronic effects on brain function of monkeys", Pharmacology of Marihuana , M. C. Brande and S. Szara, eds. (New York, Raven Press, 1976), pp. 345-356.
007J. W. Harper, R. G. Heath and W. A. Myers, "Effects of Cannabis sativa on ultrastructure of the synapse in monkey brain", Journal of Neuroscience Research , vol. 3, 1977, pp. 87-93.
008R.G. Heath and others, "Chronic marihuana smoking: its effects on function and structure of the primate brain in marihuana and membranes", in Quantitation, Metabolism, Cellular Responses, Reproduction and Brain , G. G. Nahas and W. D. Paten, eds. (New York, Pergamon Press, in press).
009C. E. Turner and others, "Constituents of Cannabis sativa L. VII: Use of silyl derivates in routine analysis", Journal of Pharmaceutical Sciences , vol. 63, No. 12 (1974), pp. 1872-1876.
010C. E. Turner and others, "Constituents of Cannabis sativa L. VIII: Possible biological application of a new method to separate cannabidiol and cannabichromene", Journal of Pharmaceutical Sciences , vol. 64, No. 5 (1975), pp. 810-814.
011C. E. Turner, K.W. Hadley and K.H. Davis, Jr., "Constituents of Cannabis sativa L . V: Stability of an analytical sample extracted with chloroform", Acta Pharmaceutica Jugoslavica , vol. 23, No. 2 (1973), pp. 89-94.
012J. H. Holley, K. W. Hadley and C. E. Turner, "Constituents of Cannabis sativa L . XI: Cannabidiol and cannabichromene in samples of known geological origin", Journal of Pharmaceutical Sciences , vol. 64, No. 5 (1975), pp. 892-895.
013I. G. Karniol and E. A. Carlini, "The content of (-)Δ 9-trans-tetrahydrocannabinol (Δ 9-THC) does not explain all biological activity of some Brazilian marihuana samples", Journal of Pharmacy and Pharmacology , vol. 24, No. 10 (1972), pp. 833-834.
014I. G. Karniol and E. A. Carlini, "Pharmacological interaction between cannabidiol and Δ 9-tetrahydrocannabinol", Psychopharmacologia, vol. 33, 1973, pp. 33-70.
015I.G. Karniol and others, "Cannabidiol interferes with the effects of Δ 9-tetrahydrocannabinol in man", European Journal of Pharmacology , vol. 28, 1974, pp. 172-177.
016I.G. Karniol and others, "Effects of Δ 9-tetrahydrocannabinol and cannabinol in man", Pharmacology, vol. 13, No. 6 (1975), pp. 502-512.
017C.E. Turner and J. Hadley, "Constituents of Cannabis sativa L. II: Absence of cannabidiol in an African variant", Journal of Pharmaceutical Sciences , vol. 62, No. 2 (1972), pp. 251-255.
018M. A. Elsohly, E.G. Boeren and C.E. Turner, "Constituents of Cannabis sativa L . An improved method for the synthesis of dl-Cannabichromene", Journal of Heterocyclic Chemistry , vol. 15, 1978, PP. 699-700.
019M. A. Elsohly, F. S. El-Feraly and C. E. Turner, "Isolation and characterization of (+)-cannabitriol and (-)-10-ethoxy-9-hydroxy- 6a( 10 a)-tetrahydrocannabinol: two new cannabinoids from Cannabis sativa L. extract", Lloydia, vol. 40, No. 3 (1977), pp. 275-280.
020E. G. Boeren, M. A. Elsohly and C. E. Turner, "Cannabiripsol: A novel Cannabis constituent", Experientia, in press.
021C. T. Turner and M. A. Elsohly, "Constituents of Cannabis sativa L. XVI: A possible decomposition pathway of Δ 9-tetrahydrocannabinol to cannabinol", Journal of Heterocyclic Chemistry , in press.
022S. Agurell and K. Leander, "Stability, transfer and absorption of cannabinoid constituents of Cannabis (Hashish) during smoking", Acta Pharmaceutical Sueccica , vol. 8, 1971, pp. 391-402.
023S. J. Gross and others, "Metabolites measured by a radioimmune technique", Nature, vol. 252, 1974, pp. 581-582.
024J. D. Teal and others, "Radioimmunoassay of cannabinoids in blood in urine", The Lancet , 1974, pp. 553-555.
025S. Agurell and others, "Quantitation of Δ 9-THC in plasma from Cannabis smokers", Pharmacy Pharmacology , vol. 25, 1973, pp. 554-558.
026C.E. Cook and others, "Radioimmunoassay of Δ 9-tetrahydrocannabinol", Cannabinoid. Assays in Humans , NIDA Research Monograph 7, R. E. Willette, ed. (United States Department of Health, Education and Welfare, 1976), pp. 15-27.
027A. R. Chase and others, "Quantitation of cannabinoids in biological fluids by radioimmunoassay", Cannabinoid Assays in Humans , NIDA Research Monograph 7, R. E. Willette, ed. (United States Department of Health, Education and Welfare, 1976), pp. 1-9.
028S. J. Cross and J. R. Soares, "Separate radioimmune measurements of body fluid Δ 9-THC and 1 l-nor-9-carboxy-Δ 9-THC", Cannabinoid Assays in Humans , NIDA Research Monograph 7, R. E. Willette, ed. (United States Department of Health, Education and Welfare, 1976), pp. 10-14.
029R. Detrick and R. Foltz, "Quantitation of Δ 9-tetrahydrocannabinol in body fluids by gas-chromatography/chemical ionization-mass spectrometry", Cannabinoid Assays in Humans , NIDA Research Monograph 7, R. E. Willette, ed. (United States Department of Health, Education and Welfare, 1976), pp. 88-95.

